



Multiple molecular and cellular mechanisms of the antitumour effect of dihydromyricetin (Review)
- Authors:
- Published online on: March 26, 2024 https://doi.org/10.3892/br.2024.1769
- Article Number: 82
-
Copyright: © Xia et al. This is an open access article distributed under the terms of Creative Commons Attribution License.
Abstract
1. Introduction
Cancer is the second most deadly disease worldwide after cardiovascular disease and one of the major threats to human health (1). According to recent estimates, the global cancer burden is expected to increase significantly in the coming decades (by 47% from 2023 to 2040) (2,3). Due to the various limitations regarding conventional treatment regimens, a growing number of researchers around the world are working to discover new anticancer agents and develop new effective modalities for cancer treatment (4). The vast majority of currently clinically approved chemotherapeutic agents are derived from a variety of natural sources, including microbes and terrestrial and marine plants (5). Commonly used anticancer drugs such as pergolide, vincristine, paclitaxel and podophyllotoxin are found in plants (6,7). Inspired by the recent success in drug development, an increasing number of research groups worldwide are working to isolate new structural leads from different plant species and to assess their potential anticancer activity.
Dihydromyricetin (DHM, PubChem compound ID: 161557), also known as ampelopsin, is a polyphenolic hydroxy-dihydroflavonol compound (8). It was first isolated from the plant Ampelopsis meliaefolia Kudo in 1940(9); it is part of a class of active natural compounds widely present in a variety of plant families, including the Umbelliferae, Asteraceae, Leguminosae, Rutaceae, Mulberry, Mucuna pruriens and Thymus families (10). DHM has a wide range of pharmacological activities, such as anti-inflammatory, anticoagulant, antibacterial, antifungal, antiviral, anticancer and antihypertensive activities (11,12). DHM has poor water solubility and is only stable at low temperatures and in a weakly acidic environment (pH 6.0) (8,13). To improve the medicinal properties (for example, the solubility, permeability and stability) of DHM, researchers have developed numerous new formulations of DHM, such as nano-formulations (14), microemulsions (15), gels (16) and crystals (17).
Several preclinical studies have revealed that DHM can inhibit the growth of a variety of cancer cells by modulating various cellular signalling pathways; for example, DHM can protect the cardiovascular system through the PI3K/Akt, Nrf2/HO-1 and SIRT3 signalling pathways (18-20) and can prevent or inhibit the growth of a variety of cancer cells through the ERK/Akt, AMPK/MAPK/XAF1, Akt-mTOR, ROS/NF-κB and mitochondrial apoptosis signalling pathways to prevent or inhibit hepatocellular carcinoma, lung cancer, colorectal cancer, breast cancer, leukaemia and melanoma (21-23). DHM can also prevent or inhibit neurodegenerative diseases through the TRL4/NF-κB and p53/p21 signalling pathways (24,25). In addition, DHM has anti-inflammatory, antibacterial, antiviral and skin-protective effects, thus it has great potential for the treatment of tumours (10). Previous studies have revealed that DHM can inhibit the proliferation of various types of tumour cells, induce apoptosis, and inhibit migration, invasion, and metastasis (12). In addition, treatment of malignant tumour cells with DHM has been shown to suppress drug resistance and increase the response to standard anticancer drugs such as adriamycin or doxorubicin (DOX), opening new avenues for cancer treatment (18,26). Therefore, in the present review, the multiple molecular and cellular mechanisms underlying the tumour-suppressive effects of DHM will be described in depth to provide ideas for the research and development of clinical treatments involving DHM.
Given that DHM has drawn widespread concern in recent years, its anticancer effects have been widely described (9,12,27), while there are few reviews that summarize the antitumour effects of DHM from the molecular mechanism level, and the reviews about the role of DHM in conventional antitumour drugs are less understood. In 2018 Zhang et al (28) suggested that DHM may be associated with several different molecules involved in cellular apoptosis, oxidative stress and inflammation. However, further research on DHM discovered that DHM is also associated with tumor cell proliferation (29), invasion and metastasis (30), and autophagy (31). To improve understanding of the antitumour mechanism of DHM, in the present review, four types of antitumour effects of DHM were summarized: Inhibition of tumour cell proliferation, promotion of apoptosis, inhibition of tumour cell invasion and migration, scavenging of tumour cell reactive oxygen species (ROS), and induction of autophagy. Relevant articles were screened from recent years and the results related to the molecular and cellular mechanisms were summarized. The present review also summarized the potential application of DHM in combination with conventional antitumour drugs, as well as the advantages and disadvantages of DHM as an antitumour drug.
2. Multiple molecular and cellular mechanisms of the antitumour activity of DHM
DHM induces tumour cell cycle arrest and inhibits cell proliferation
The process of the cell cycle is divided into interphase (G1 phase, S phase, G2 phase), division phase (M phase), and stationary phase (G0 phase), and the normal progression of the cell cycle is regulated by various mechanisms of action to ensure orderly cell division (32); if any of the related factors are dysregulated, the cell cycle is terminated (32,33). The central cell cycle regulation mechanism is the Cyclin-CDK-CDI mechanism, and CDK inhibitor (CKI) genes have positive regulators such as cyclin and p16 and negative regulators such as p53 and p21 (34,35). The overexpression of positive regulators or the absence of negative regulators results in the downregulation of the cell cycle threshold and weakening of cell cycle function, resulting in reduced sensitivity of cells to exogenous regulatory signals (32,35). CDC genes (cell division cycle genes) are a class of genes whose expression is cell cycle-dependent or directly involved in cell cycle regulation; these genes mainly include Cyclin genes, CDK genes and CKI genes (36,37). In addition, DNA polymerase and DNA ligase genes related to DNA replication are also part of the CDC gene family (38).
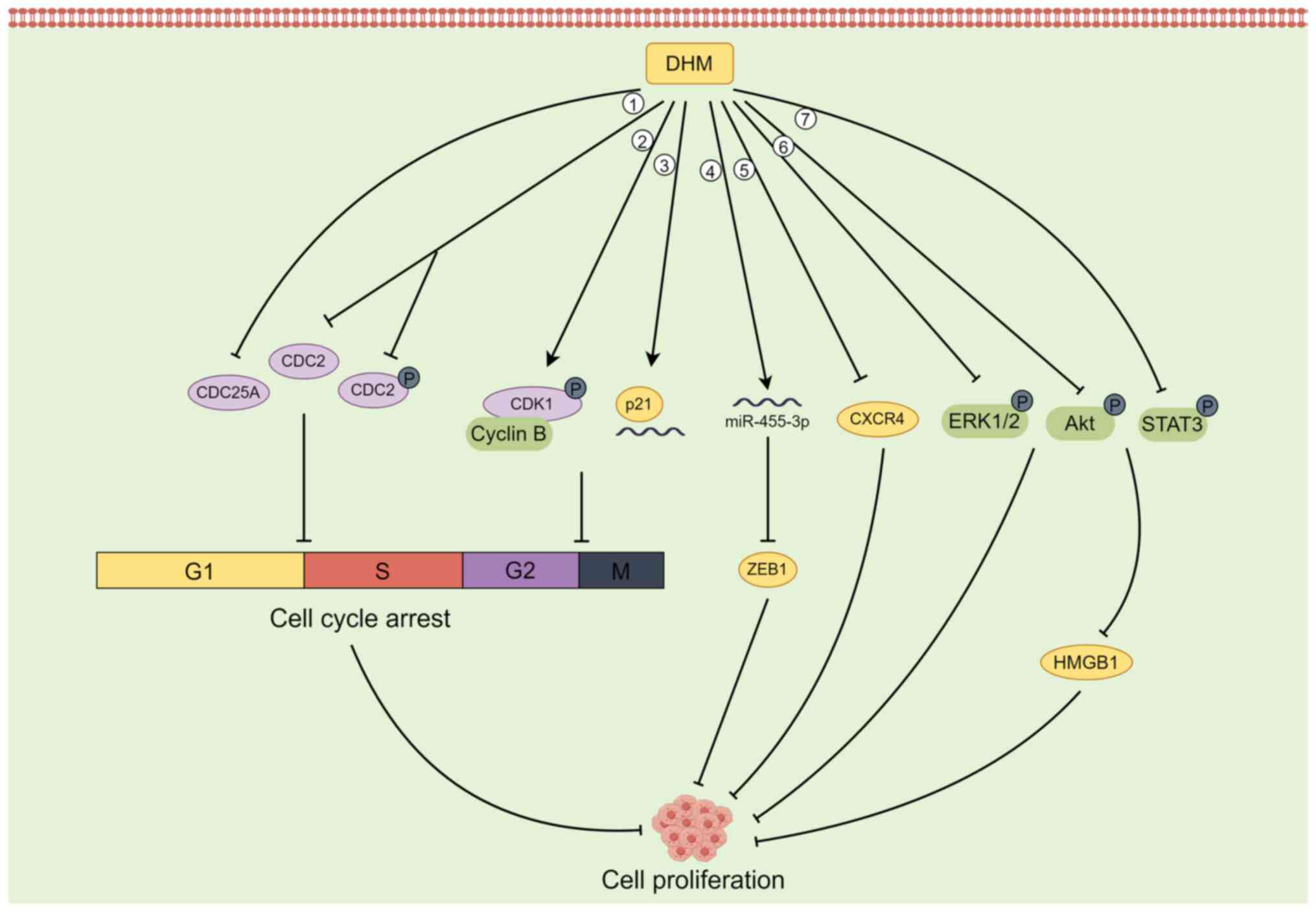
Several studies have reported that in some tumour cells, DHM can inhibit cell proliferation by blocking the G2/M phase and G1/S phase of the cell cycle (29,39,40). For example, DHM can inactivate the CDK1/cell cycle protein B1 complex by phosphorylating CDK1, thus inducing G2/M phase cell cycle arrest and inhibiting the growth of hepatocellular carcinoma cells (29). In SK-MEL-28 melanoma cells, DHM downregulates the expression of the CDC25A, CDC2, and phosphorylated (p-)CDC2 proteins to induce cell cycle arrest in the G1/S phase (39). DHM also induces G2-M arrest of the U2OS cell cycle in osteosarcoma cells by increasing the levels of p21 protein and RNA and inhibiting cell proliferation (40).
In addition to inhibiting the proliferation of tumour cells by blocking the cell cycle, DHM can also target the activation of ERK1/2 and Akt to inhibit the proliferative potential of fibroblasts in lung cancer cells (41) and upregulate the expression of p53 to inhibit the proliferation of hepatocellular carcinoma cells (42). In animal studies using an in situ prostate tumour model, Ni et al (43) reported that DHM inhibited the proliferation of PC-3 tumours in a dose-dependent manner, which was associated with a decrease in the CXCR4 protein. In BGC-823 gastric cancer cells, DHM induced a decrease in the expression of Cyclin D1, Cyclin E1 and N-cadherin; increased the expression of E-cadherin; inhibited the phosphorylation of Akt and STAT3; and downregulated the expression of HMGB1 in cells. Therefore, DHM inhibits BGC-823 cell proliferation and migration by regulating the activation of the Akt/STAT3 signalling pathway and the expression of HMGB1(44). In cholangiocarcinoma (CAA) cells, DHM upregulated the expression of miR-455-3p, which in turn inhibited the expression of its downstream target ZEB1, thereby suppressing the proliferation of CAA cells (45). ZEB1 is a transcription factor that promotes metastasis and stem cell characteristics, and it is aberrantly expressed in CAA cells (46) (Fig. 1).
DHM induces the apoptosis of tumour cells and affects signalling pathways
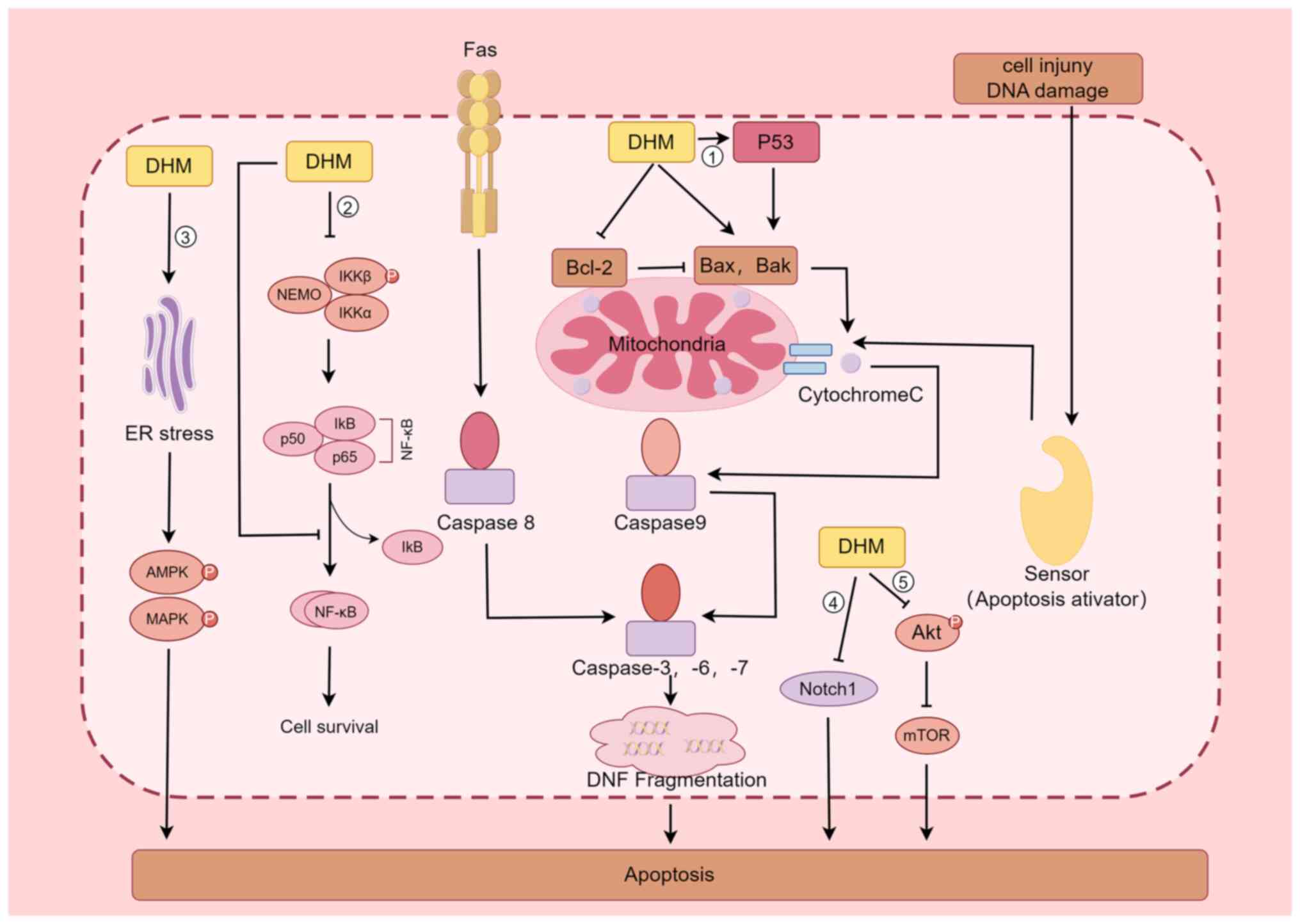
The apoptotic pathway includes both exogenous and endogenous pathways, the former being mediated by receptors such as TNF-α and FAS-L, while the latter uses mitochondria as the main site of apoptosis induction by regulating the permeability of their outer membrane (47). When various stress responses, DNA damage and abnormal cell signalling occur, Bax is activated, and under the regulation of this protein, the permeability of the mitochondrial outer membrane increases, and Cytochrome C is released into the cytoplasm, where it binds to Apaf-1 in the presence of dATP, causing Apaf-1 to expose its Card structural domain and bind to the Card of Procaspase-9 to form an apoptotic complex; this complex in turn activates Caspase-9, which further activates Caspase-3, thus causing apoptosis (48-50). By contrast, Bcl-2 plays the opposite role as Bax in inhibiting Cytochrome C release, thus inhibiting the process of apoptosis; therefore, regulation of the expression of Bcl-2 has an important impact on the apoptosis of tumour cells (51). Because of the inverse effects of Bcl-2 and Bax, when cells are stimulated, the ratio of the two determines cell survival (52). Liu et al (42) discovered that DHM significantly increases p53 protein expression in four types of hepatocellular carcinoma cells (HepG2, QGY7701, Hepal-6, and MHcc97L), which activated Bax and Bak, and inhibited Bcl-2 expression, which in turn activated Caspase-3 and eventually led to the apoptosis of tumour cells; the regulatory effect was dose-dependent. Ji et al (53) revealed that DHM significantly (P<0.05) inhibited AGS cell proliferation and induced cell cytotoxicity in a dose- and time-dependent manner; DHM also regulated the expression of apoptosis-related genes such as p53 and Bcl-2 in a dose- and time-dependent manner in AGS cells treated with DHM, as determined by western blotting (53).
In addition to inducing the apoptosis in tumour cells through the p53-mediated signalling pathway, DHM is associated with the Nuclear Factor Kappa B Subunit 1(NF-κB) signalling pathway (22,23,54). Han et al (54) reported that in the leukemic cell lines HL60 and K562, DHM induced apoptosis through nuclear condensation, induced loss of mitochondrial membrane potential, increased ROS production, activated Caspase-9, Caspase-3 and poly ADP-ribose polymerase, and regulated the expression of Bcl-2 family members; these authors also reported that DHM induced apoptosis in the leukaemia cell lines HL60 and K562, possibly through the NF-κB signalling pathway. Li et al (22) examined the expression of phosphorylated nuclear factor kappa B kinase subunit β (p-IKKβ), phosphorylated nuclear factor kappa B kinase subunit α (p-IKKα), nuclear factor kappa Bα (IκB-α) inhibitor, and NF-κB/p65 in a nasopharyngeal carcinoma CNE-2 cell line via western blot analysis and confocal laser scanning microscopy; the observed nuclear translocation of NF-κB/p65 indicated that DHM promotes the inactivation of p-IKKβ and p-IKKα and blocks the nuclear translocation of the NF-κB subunit p65, promoting the apoptosis of CNE-2 cells in nasopharyngeal carcinoma. Guo et al (23) treated a rat model of fatty liver disease with DHM and found that DHM inhibited the protein expression of NF-κB, p53 and Bax, acting as hepatoprotective agent.
In addition, DHM promotes the apoptosis of tumour cells through several other pathways (55-59). For example, DHM inhibits the activation of Akt, which in turn inhibits the formation of the mTOR complex and promotes apoptosis in breast cancer cells (55). DHM is a biologically active natural chemopreventive agent and a potent mTOR inhibitor that may be a useful chemotherapeutic agent for breast cancer treatment (55). Lu et al (56) revealed that Notch1 is involved in the development of HCC and that DHM inhibits cell proliferation and promotes apoptosis by downregulating Notch1 expression. A study by Ye et al (57) demonstrated that in cisplatin-resistant nasopharyngeal carcinoma cell lines (Hone1/Cis and CNE1/Cis), cotreatment with DHM increased the growth-inhibitory effect of cisplatin by blocking the Wnt/β-catenin signalling pathway to increase the antitumour activity of cisplatin in nasopharyngeal carcinoma. In a mouse model of cerebral ischaemia-reperfusion injury, DHM inhibited oxidative stress and apoptosis in mouse hippocampal neuronal HT22 cells by activating the Nrf2/HO-1 signalling pathway (58). Treatment of colon cancer cells with DHM resulted in dose- and time-dependent apoptosis through the activation of endoplasmic reticulum (ER) stress, 5'-adenosine monophosphate-activated protein kinase (AMPK) and JNK/p38 MAPKs, and AMPK/MAPK/XAF1 signalling initiated by DHM through the ER stress pathway, which induces apoptosis in colon cancer cells (59) (Fig. 2).
The mechanisms by which DHM inhibits tumour cell invasion and metastasis
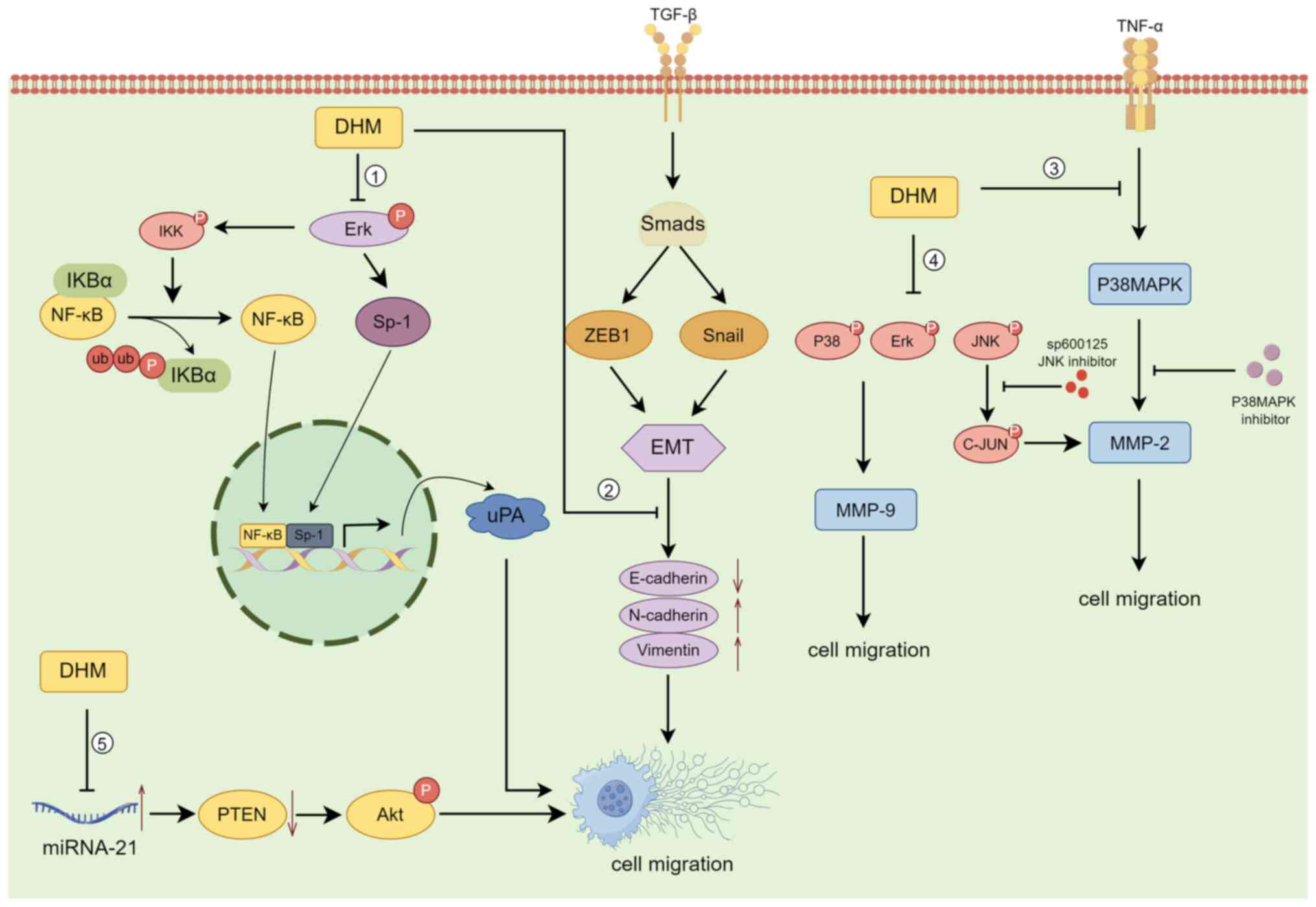
The invasion-migration cascade is a complex biological process that includes the following major events: (i) Cell migration and local invasion of the basement membrane, (ii) invasion of the vasculature and/or lymphatic system, (iii) survival in the circulation, (iv) arrest and extravasation at distant organ sites, and (v) colonization at metastatic sites (60,61). According to previous case reports, >80% of cancer patients succumb to tumour invasion/metastasis, making it one of the major causes of death in cancer patients. The decrease in 5-year survival upon metastasis is particularly severe in patients with osteosarcoma (62). DHM can suppress the invasion and metastasis of osteosarcoma cells by blocking the TNF-α/p38MAPK/MMP-2 signalling pathway (30). Moreover, Chou et al (63) reported that DHM regulates osteosarcoma metastasis through the ERK pathway; it also inhibits metastasis by suppressing the expression of the downstream urokinase plasminogen activator through the inhibition of SP-1 and NF-κB.
DHM inhibits the invasion and migration of human retinal pigment epithelial cells (ARPE-19) by decreasing the expression of MMP-2(64). By contrast, Zhang et al (65) reported that DHM significantly inhibited the migration and invasion of SK-Hep-1 and MHCC97L hepatocellular carcinoma cells by decreasing MMP-9 protein expression, while downregulation of MMP-9 protein expression was closely associated with increased PKC-δ protein levels and decreased phosphorylation of p38, ERK1/2 and JNK in SK-Hep-1 and MHCC97L cells. In the ovarian cancer cell line A2780, DHM upregulated E-cadherin and downregulated N-cadherin and Vimentin in the Snail signalling pathway in a concentration- and time-dependent manner, inhibiting the nuclear translocation of NF-κB; these results suggested that DHM inhibits epithelial-mesenchymal transition (EMT) via the NF-κB/Snail pathway and suppresses ovarian cancer cell invasion (66). DHM also was also demonstrated to inhibit the migration and invasion of human gastric cancer MKN45 cells and reverse EMT through the downregulation of MMP-2 expression via the JNK signalling pathway (67). In an in vivo experiment on mice transplanted with B16 melanoma cells, Zheng et al (68) reported that mice given DHM at doses of 150, 200 and 250 mg/kg exhibited a significant reduction in the number of metastatic tumours compared with those in the control group, demonstrating the anti-invasive and anti-metastatic effects of DHM on B16 melanoma. Moreover, DHM inhibits the migration and invasion of nasopharyngeal carcinoma cells by suppressing the ERK1/2 signalling pathway and suppressing MMP-2 expression (69); it also inhibits the proliferation, migration and invasion of CAA HCCC9810 and TFK-1 cells by regulating miR-21 and promoting apoptosis (70) (Fig. 3).
DHM regulates intracellular ROS levels and induces autophagy in tumour cells
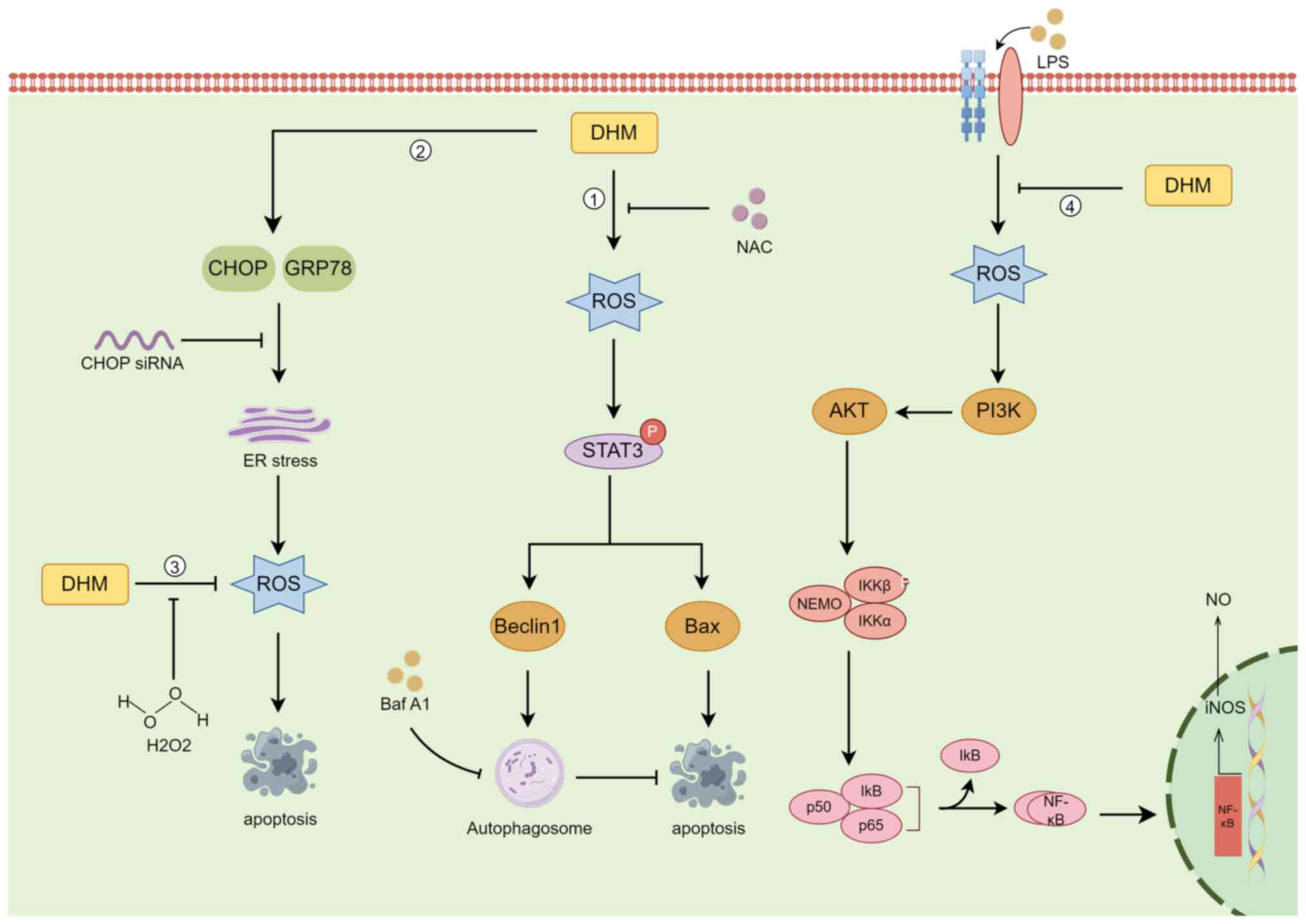
ROS are highly reactive substances containing oxygen radicals, and ROS are produced in various biochemical reactions in cellular organelles, such as the ER, mitochondria and peroxisomes, as a by-product of normal oxygen metabolism (71). ROS are cytotoxic molecules that stimulate apoptosis, but high levels of ROS can induce tumorigenesis, leading to uncontrolled cancer cell proliferation (72). Autophagy is an evolutionarily conserved catabolic mechanism by which eukaryotic cells recycle or degrade internal components through a membrane transport pathway (73). The process of autophagy is divided into four key steps: Initiation, nucleation, maturation and degradation (74). In cancer, autophagy plays a dichotomous role, that is, it inhibits tumorigenesis and supports tumour development (75).
DHM induces autophagy in head and neck squamous cell carcinoma (HNSCC) through the phosphorylation and activation of the STAT3 transcription factor (31). In HNSCC cells, the increase in ROS levels was proportional to the increase in DHM concentration, and DHM activated p-STAT3-dependent autophagy through ROS production in HNSCC (31). Zhou et al (76) demonstrated that in the human breast cancer cell lines MCF-7 and MDA-MB-231, DHM had antitumour effects through ROS production, and the ER stress pathway had antitumour effects on breast cancer cells. However, in human melanoma cells, autophagy has a protective role in DHM-induced apoptosis, and pharmacological inhibition or genetic blockade of autophagy increases DHM-induced cell death and apoptosis (77). Liu et al (78) evaluated the effects of DHM on the induction of ROS accumulation and activation of mitochondrial signalling pathways in human hepatocellular carcinoma HepG2 cells. DHM reduced the accumulation of ROS in a concentration-dependent manner, while the expression of proteins involved in the apoptotic program increased in a concentration-dependent manner. This suggests that ROS can act as redox signalling messengers that regulate DHM-induced apoptosis (78). In a pharmacobiochemical study of the effect of DHM on inflammation in a RAW264.7 macrophage model, DHM was discovered to inhibit the accumulation of ROS, suppress the release of NO and the proinflammatory cytokines IL-1β, IL-6 and TNF-α, and suppress lipopolysaccharide-induced inducible nitric oxide synthase by inhibiting nuclear factor-κB (NF-κB) activation (79). In cutaneous squamous cell carcinoma (CSCC), DHM induced TFEB (Ser142) dephosphorylation, activated TFEB nuclear translocation, increased TFEB reporter activity, decreased lncRNA MALAT1 expression, and induced CSCC cell death by inducing excessive autophagy via the MALAT1-TFEB pathway (80) (Fig. 4).
3. DHM attenuates the drug resistance of tumour cells and increases their sensitivity to chemotherapeutic drugs
DHM is mainly extracted from the plant Garcinia cambogia and is an antitumour drug with high efficiency, low toxicity and few side effects (81). The focus of present review was mainly to explore its antitumour mechanism in combination with existing chemotherapeutic drugs (82-84). Several studies have demonstrated that the combination of DHM has a stronger antitumour effect than single drugs (85-91). Jiang et al (85) reported that DHM increases the chemosensitivity of hepatocellular carcinoma cell lines to nedaplatin (NDP) through the p53/Bcl-2 pathway while reducing the damage caused by NDP to normal hepatocytes and thus protecting normal hepatocytes. Another study revealed that DHM increases the chemosensitivity of leukemic NB4 cells to all-trans retinoic acid through modulation of the p38-STAT1 signalling pathway, thereby further inhibiting cancer cell growth (86). Wang et al (87) reported that DHM increased the chemosensitivity of CRC cells to oxaliplatin (OXA). DHM increased the chemosensitivity to OXA, promoted OXA-induced apoptosis, and suppressed the accumulation of 5(6)-carboxy-2,7-dichlorofluorescein in OXA-resistant HCT116/L-OHP CRC cells. DHM inhibited the growth of colorectal cancer cells by suppressing MRP2 expression and activity in the HCT116/OXA and HCT8/vincristine (VCR) colorectal cancer cell lines; these changes in MRP2 expression and promoter activity restored the chemosensitivity of these two cell lines to OXA and VCR (88). In a mouse model of AOM/DSS-induced colorectal cancer, DHM increased the therapeutic effect of irinotecan (CPT-11) (89).
Ovarian cancer is one of the leading causes of cancer-related death in gynaecologic malignancies, and resistance to chemotherapeutic agents remains a major challenge in ovarian cancer treatment (90). Xu et al (90) reported that DHM significantly increased the sensitivity of ovarian cancer cells to paclitaxel and DOX by inhibiting the expression of survivin, a member of the IAP family of apoptosis-inhibitory proteins. One of the most common treatments for gastric cancer is chemotherapy, but multidrug resistance often leads to the failure of anticancer therapy, and the combination of DHM and mitomycin increases the inhibitory effect of mitomycin on the proliferation of gastric cancer cells (91).
4. Advantages and challenges of DHM as an antitumour drug
The advantages of DHM as an antitumour drug for combination with chemotherapy are as follows: It has fewer toxic side effects than chemotherapy, it has multiple antitumour mechanisms, and it has a lower risk of drug resistance than chemotherapy alone (81-84). Several studies confirmed that DHM was not cytotoxic to the immortalized normal human hepatocyte line LO-2(92), the normal prostate epithelial cell line PrEC (43), or the normal mammary epithelial cell line MCF-10A (76). Moreover, Dong et al (93) reported that DHM pre-treatment regulated APAP metabolism by regulating the expression of UDP-glucuronosyltransferase 1 and cytochrome P4502E1 to ameliorate APAP-associated hepatocyte necrosis and stimulate liver regeneration. DHM can be used in combination with a variety of known chemotherapeutic agents, such as DHM in combination with NDP, to regulate the balance of Bcl-2/Bax and Bcl-2/Bak ratios through the p53/Bcl-2 signalling pathway and inhibit NDP-induced ROS production, thereby increasing the chemosensitivity of hepatocellular carcinoma cells to NDP (85). DHM increases the antitumour activity of adriamycin (ADR) and prevents ADR-induced DIC in a p53-dependent manner by inhibiting MDM2-mediated degradation of ARC via ubiquitination (90).
Although DHM is a promising cancer treatment, its chemical instability and low bioavailability hinder its application (9). The phenolic hydroxyl structure of DHM makes it unstable (93). In particular, when DHM is exposed to light, pH buffers, pepsin and trypsin, it undergoes various chemical reactions, such as oxidation, hydrolysis, cleavage, reduction and decomposition, to produce metabolites (93). Pharmacokinetic studies have also shown that DHM is not readily absorbed into the bloodstream and is unstable in the intestinal environment (94). Therefore, the combination of DHM with other chemotherapeutic drugs requires consideration of differences in the physicochemical properties, absorption sites, pharmacokinetic behaviours, and effective doses of different drugs (95).
5. Conclusions and perspectives
The present review mainly summarized the molecular mechanism underlying the inhibitory effect of DHM on tumours. A total of four effects of DHM were described: It inhibits tumour cell proliferation, promotes apoptosis, inhibits invasion and migration, clears ROS, and induces autophagy. The effects of DHM were also summarized in combination with several traditional antitumour drugs, as well as the advantages and disadvantages of DHM as an antitumour drug. Currently, there are two reported clinical trials related to DHM: In a double-blind clinical trial, Chen et al (96) conducted a three-month follow-up observation on 60 adult patients with non-alcoholic fatty liver disease and it was identified that the serum levels of alanine, aspartate aminotransferase, γ-glutamyl transpeptidase, glucose, low-density lipoprotein-cholesterol and apolipoprotein B, and the homeostasis model assessment of insulin resistance index were significantly decreased in the DHM group compared with the placebo group, and DHM supplementation improves glucose and lipid metabolism as well as various biochemical parameters in patients with non-alcoholic fatty liver disease. Ran et al (97) conducted a follow-up experiment on 80 participants with type 2 diabetes mellitus (T2DM) and it was revealed that compared with the placebo group, the levels of fasting blood glucose, glycosylated albumin, cystatin C and retinol binding protein-4 in the DHM group significantly decreased, and DHM can effectively improve the blood sugar control in patients with T2DM. However, there are no studies reporting that DHM has been tested in any clinical trials for treating cancer. The antitumour mechanisms of DHM are diverse and not limited to these four aspects, and the antitumour effects of DHM on different tumour cells may be the result of a combination of mechanisms.
With advances in diagnostic and treatment technology, the mortality rate of cancer patients is gradually decreasing. However, innovative and more effective drugs are needed to further suppress the progression of cancer. Natural herbal medicines are attracting increasing attention because they offer clear advantages in terms of research and development and medical costs. DHM, a flavonoid extracted from the stem and leaves of buttercups, has promising anticancer effects. In the present review, different molecular and cellular mechanisms by which DHM induces antitumour effects were described. DHM has been demonstrated to be highly effective and to have low toxicity and few side effects. The combination of DHM with other existing anticancer drugs could increase the inhibitory effects on tumour cells. Thus, the present review provided a comprehensive reference for the development of DHM as an anticancer drug.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
RZ and TX designed the study. TX collected the information and wrote the manuscript. RZ was responsible for handling the revisions. All authors contributed to the article and approved the submitted version. Data authentication is not applicable. All the authors have read and approved the final manuscript.
Ethics approval and consent to participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
|
Mullard A: Addressing cancer's grand challenges. Nat Rev Drug Discov. 19:825–826. 2020.PubMed/NCBI View Article : Google Scholar | |
|
Kashyap D, Tuli HS, Yerer MB, Sharma A, Sak K, Srivastava S, Pandey A, Garg VK, Sethi G and Bishayee A: Natural product-based nanoformulations for cancer therapy: Opportunities and challenges. Semin Cancer Biol. 69:5–23. 2021.PubMed/NCBI View Article : Google Scholar | |
|
Chang L, Ruiz P, Ito T and Sellers WR: Targeting pan-essential genes in cancer: Challenges and opportunities. Cancer Cell. 39:466–479. 2021.PubMed/NCBI View Article : Google Scholar | |
|
Shafabakhsh R and Asemi Z: Quercetin: A natural compound for ovarian cancer treatment. J Ovarian Res. 12(55)2019.PubMed/NCBI View Article : Google Scholar | |
|
Kim C and Kim B: Anti-Cancer natural products and their bioactive compounds inducing ER stress-mediated apoptosis: A review. Nutrients. 10(1021)2018.PubMed/NCBI View Article : Google Scholar | |
|
Demain AL and Vaishnav P: Natural products for cancer chemotherapy. Microb Biotechnol. 4:687–699. 2011.PubMed/NCBI View Article : Google Scholar | |
|
Zuo W and Kwok HF: Development of marine-derived compounds for cancer therapy. Mar Drugs. 19(342)2021.PubMed/NCBI View Article : Google Scholar | |
|
Sun CC, Li Y, Yin ZP and Zhang QF: Physicochemical properties of dihydromyricetin and the effects of ascorbic acid on its stability and bioavailability. J Sci Food Agric. 101:3862–3869. 2021.PubMed/NCBI View Article : Google Scholar | |
|
Wu J, Xiao Z, Li H, Zhu N, Gu J, Wang W, Liu C, Wang W and Qin L: Present status, challenges, and prospects of dihydromyricetin in the battle against cancer. Cancers (Basel). 14(3487)2022.PubMed/NCBI View Article : Google Scholar | |
|
Sun Y, Liu S, Yang S, Chen C, Yang Y, Lin M, Liu C, Wang W, Zhou X, Ai Q, et al: Mechanism of dihydromyricetin on inflammatory diseases. Front Pharmacol. 12(794563)2022.PubMed/NCBI View Article : Google Scholar | |
|
Cheng P, Gui C, Huang J, Xia Y, Fang Y, Da G and Zhang X: Molecular mechanisms of ampelopsin from Ampelopsis megalophylla induces apoptosis in HeLa cells. Oncol Lett. 14:2691–2698. 2017.PubMed/NCBI View Article : Google Scholar | |
|
Chen L, Shi M, Lv C, Song Y, Wu Y, Liu S, Zheng Z, Lu X and Qin S: Dihydromyricetin acts as a potential redox balance mediator in cancer chemoprevention. Mediators Inflamm. 2021(6692579)2021.PubMed/NCBI View Article : Google Scholar | |
|
Sun J, Wang Y, Tang W and Gong J: Enantioselectivity of chiral dihydromyricetin in multicomponent solid solutions regulated by subtle structural mutation. IUCrJ. 10(Pt 2):164–176. 2023.PubMed/NCBI View Article : Google Scholar | |
|
Geng S, Yuan Y, Jiang X, Zhang R, Ma H, Liang G and Liu B: An investigation on pickering nano-emulsions stabilized by dihydromyricetin/high-amylose corn starch composite particles: Preparation conditions and carrier properties. Curr Res Food Sci. 6(100458)2023.PubMed/NCBI View Article : Google Scholar | |
|
Lyu Q, Chen L, Lin S, Cao H and Teng H: A designed self-microemulsion delivery system for dihydromyricetin and its dietary intervention effect on high-fat-diet fed mice. Food Chem. 390(132954)2022.PubMed/NCBI View Article : Google Scholar | |
|
Ye J, Bao S, Zhao S, Zhu Y, Ren Q, Li R, Xu X and Zhang Q: Self-Assembled micelles improve the oral bioavailability of dihydromyricetin and anti-acute alcoholism activity. AAPS PharmSciTech. 22(111)2021.PubMed/NCBI View Article : Google Scholar | |
|
Liu L, Li Y, Zhang M, Zhang Y and Lou B: A Drug-Drug cocrystal of dihydromyricetin and pentoxifylline. J Pharm Sci. 111:82–87. 2022.PubMed/NCBI View Article : Google Scholar | |
|
Sun Z, Lu W, Lin N, Lin H, Zhang J, Ni T, Meng L, Zhang C and Guo H: Dihydromyricetin alleviates doxorubicin-induced cardiotoxicity by inhibiting NLRP3 inflammasome through activation of SIRT1. Biochem Pharmacol. 175(113888)2020.PubMed/NCBI View Article : Google Scholar | |
|
Wei C, Chen X, Chen D, Yu B, Zheng P, He J, Chen H, Yan H, Luo Y and Huang Z: Dihydromyricetin enhances intestinal antioxidant capacity of growing-finishing pigs by activating ERK/Nrf2/HO-1 signaling pathway. Antioxidants (Basel). 11(704)2022.PubMed/NCBI View Article : Google Scholar | |
|
Zhou J, Hou P, Yao Y, Yue J, Zhang Q, Yi L and Mi M: Dihydromyricetin improves high-fat diet-induced hyperglycemia through ILC3 Activation via a SIRT3-Dependent Mechanism. Mol Nutr Food Res. 66(e2101093)2022.PubMed/NCBI View Article : Google Scholar | |
|
Zhang ZY, Liu C, Wang PX, Han YW, Zhang YW, Hao ML, Song ZX and Zhang XY: Dihydromyricetin Alleviates H9C2 cell apoptosis and autophagy by regulating CircHIPK3 Expression and PI3K/AKT/mTOR pathway. Chin J Integr Med. 29:434–440. 2023.PubMed/NCBI View Article : Google Scholar | |
|
Li CH, Ding H, Shi JL, Huang B, Ding H, Lin HG, Zeng JC, Zhao Y and Luo GQ: Dihydromyricetin promotes apoptosis, suppresses proliferation and tumor necrosis factor-α-mediated nuclear factor kappa-B activation in nasopharyngeal carcinoma CNE-2 cell. J Tradit Chin Med. 41:367–375. 2021.PubMed/NCBI View Article : Google Scholar | |
|
Guo L, Zhang H and Yan X: Protective effect of dihydromyricetin revents fatty liver through nuclear factor-κB/p53/B-cell lymphoma 2-associated X protein signaling pathways in a rat model. Mol Med Rep. 19:1638–1644. 2019.PubMed/NCBI View Article : Google Scholar | |
|
Jing N and Li X: Retraction on ‘Dihydromyricetin attenuates inflammation through TLR4/NF-kappa B pathway’. Open Med (Wars). 16(1082)2021.PubMed/NCBI View Article : Google Scholar | |
|
Liu M, Guo H, Li Z, Zhang C, Zhang X, Cui Q and Tian J: Molecular level insight into the benefit of myricetin and dihydromyricetin uptake in patients with Alzheimer's diseases. Front Aging Neurosci. 12(601603)2020.PubMed/NCBI View Article : Google Scholar | |
|
Zhu H, Luo P, Fu Y, Wang J, Dai J, Shao J, Yang X, Chang L, Weng Q, Yang B and He Q: Dihydromyricetin prevents cardiotoxicity and enhances anticancer activity induced by adriamycin. Oncotarget. 6:3254–3267. 2015.PubMed/NCBI View Article : Google Scholar | |
|
Wang Y, Wang J, Xiang H, Ding P, Wu T and Ji G: Recent update on application of dihydromyricetin in metabolic related diseases. Biomed Pharmacother. 148(112771)2022.PubMed/NCBI View Article : Google Scholar | |
|
Zhang J, Chen Y, Luo H, Sun L, Xu M, Yu J, Zhou Q, Meng G and Yang S: Recent update on the pharmacological effects and mechanisms of dihydromyricetin. Front Pharmacol. 9(1204)2018.PubMed/NCBI View Article : Google Scholar | |
|
Huang H, Hu M, Zhao R, Li P and Li M: Dihydromyricetin suppresses the proliferation of hepatocellular carcinoma cells by inducing G2/M arrest through the Chk1/Chk2/Cdc25C pathway. Oncol Rep. 30:2467–2475. 2013.PubMed/NCBI View Article : Google Scholar | |
|
Liu C, Zhao P, Yang Y, Xu X, Wang L and Li B: Ampelopsin suppresses TNF-α-induced migration and invasion of U2OS osteosarcoma cells. Mol Med Rep. 13:4729–4736. 2016.PubMed/NCBI View Article : Google Scholar | |
|
Fan TF, Wu TF, Bu LL, Ma SR, Li YC, Mao L, Sun ZJ and Zhang WF: Dihydromyricetin promotes autophagy and apoptosis through ROS-STAT3 signaling in head and neck squamous cell carcinoma. Oncotarget. 7:59691–59703. 2016.PubMed/NCBI View Article : Google Scholar | |
|
Martínez-Alonso D and Malumbres M: Mammalian cell cycle cyclins. Semin Cell Dev Biol. 107:28–35. 2020.PubMed/NCBI View Article : Google Scholar | |
|
Tokunaga Y, Otsuyama KI and Hayashida N: Cell cycle regulation by heat shock transcription factors. Cells. 11(203)2022.PubMed/NCBI View Article : Google Scholar | |
|
Gonzales AJ, Goldsworthy TL and Fox TR: Chemical transformation of mouse liver cells results in altered cyclin D-CDK protein complexes. Carcinogenesis. 19:1093–1102. 1998.PubMed/NCBI View Article : Google Scholar | |
|
Zhang M, Zhang L, Hei R, Li X, Cai H, Wu X, Zheng Q and Cai C: CDK inhibitors in cancer therapy, an overview of recent development. Am J Cancer Res. 11:1913–1935. 2021.PubMed/NCBI | |
|
Wood DJ and Endicott JA: Structural insights into the functional diversity of the CDK-cyclin family. Open Biol. 8(180112)2018.PubMed/NCBI View Article : Google Scholar | |
|
Riba A, Oravecz A, Durik M, Jiménez S, Alunni V, Cerciat M, Jung M, Keime C, Keyes WM and Molina N: Cell cycle gene regulation dynamics revealed by RNA velocity and deep-learning. Nat Commun. 13(2865)2022.PubMed/NCBI View Article : Google Scholar | |
|
Lee JH and Berger JM: Cell Cycle-Dependent control and roles of DNA Topoisomerase II. Genes (Basel). 10(859)2019.PubMed/NCBI View Article : Google Scholar | |
|
Zeng G, Liu J, Chen H, Liu B, Zhang Q, Li M and Zhu R: Dihydromyricetin induces cell cycle arrest and apoptosis in melanoma SK-MEL-28 cells. Oncol Rep. 31:2713–2719. 2014.PubMed/NCBI View Article : Google Scholar | |
|
Zhao Z, Yin JQ, Wu MS, Song G, Xie XB, Zou C, Tang Q, Wu Y, Lu J, Wang Y, et al: Dihydromyricetin activates AMP-activated protein kinase and P38(MAPK) exerting antitumor potential in osteosarcoma. Cancer Prev Res (Phila). 7:927–938. 2014.PubMed/NCBI View Article : Google Scholar | |
|
Fan KJ, Yang B, Liu Y, Tian XD and Wang B: Inhibition of human lung cancer proliferation through targeting stromal fibroblasts by dihydromyricetin. Mol Med Rep. 16:9758–9762. 2017.PubMed/NCBI View Article : Google Scholar | |
|
Liu J, Shu Y, Zhang Q, Liu B, Xia J, Qiu M, Miao H, Li M and Zhu R: Dihydromyricetin induces apoptosis and inhibits proliferation in hepatocellular carcinoma cells. Oncol Lett. 8:1645–1651. 2014.PubMed/NCBI View Article : Google Scholar | |
|
Ni F, Gong Y, Li L, Abdolmaleky HM and Zhou JR: Flavonoid ampelopsin inhibits the growth and metastasis of prostate cancer in vitro and in mice. PLoS One. 7(e38802)2012.PubMed/NCBI View Article : Google Scholar | |
|
Wang S, Ge F, Cai T, Qi S and Qi Z: Dihydromyricetin inhibits proliferation and migration of gastric cancer cells through regulating Akt/STAT3 signaling pathways and HMGB1 expression. Nan Fang Yi Ke Da Xue Xue Bao. 41:87–92. 2021.PubMed/NCBI View Article : Google Scholar : (In Chinese). | |
|
Li X, Yang ZS, Cai WW, Deng Y, Chen L and Tan SL: Dihydromyricetin inhibits tumor growth and epithelial-mesenchymal transition through regulating miR-455-3p in Cholangiocarcinoma. J Cancer. 12:6058–6070. 2021.PubMed/NCBI View Article : Google Scholar | |
|
Lobe C, Vallette M, Arbelaiz A, Gonzalez-Sanchez E, Izquierdo L, Pellat A, Guedj N, Louis C, Paradis V, Banales JM, et al: Zinc Finger E-Box Binding Homeobox 1 promotes cholangiocarcinoma progression through tumor dedifferentiation and tumor-stroma paracrine signaling. Hepatology. 74:3194–3212. 2021.PubMed/NCBI View Article : Google Scholar | |
|
Xu X, Lai Y and Hua ZC: Apoptosis and apoptotic body: Disease message and therapeutic target potentials. Biosci Rep. 39(BSR20180992)2019.PubMed/NCBI View Article : Google Scholar | |
|
Morana O, Wood W and Gregory CD: The apoptosis paradox in cancer. Int J Mol Sci. 23(1328)2022.PubMed/NCBI View Article : Google Scholar | |
|
Cheng X and Ferrell JE Jr: Apoptosis propagates through the cytoplasm as trigger waves. Science. 361:607–612. 2018.PubMed/NCBI View Article : Google Scholar | |
|
Rahmani M, Nkwocha J, Hawkins E, Pei X, Parker RE, Kmieciak M, Leverson JD, Sampath D, Ferreira-Gonzalez A and Grant S: Cotargeting BCL-2 and PI3K Induces BAX-Dependent mitochondrial apoptosis in AML cells. Cancer Res. 78:3075–3086. 2018.PubMed/NCBI View Article : Google Scholar | |
|
Zuo Y, Xu Q, Lu Y, Sun D, Wang K, Lei Y, Liang X and Li Y: Dihydromyricetin induces apoptosis in a human choriocarcinoma cell line. Oncol Lett. 16:4229–4234. 2018.PubMed/NCBI View Article : Google Scholar | |
|
Chipuk JE, McStay GP, Bharti A, Kuwana T, Clarke CJ, Siskind LJ, Obeid LM and Green DR: Sphingolipid metabolism cooperates with BAK and BAX to promote the mitochondrial pathway of apoptosis. Cell. 148:988–1000. 2012.PubMed/NCBI View Article : Google Scholar | |
|
Ji FJ, Tian XF, Liu XW, Fu LB, Wu YY, Fang XD and Jin HY: Dihydromyricetin induces cell apoptosis via a p53-related pathway in AGS human gastric cancer cells. Genet Mol Res. 14:15564–15571. 2015.PubMed/NCBI View Article : Google Scholar | |
|
Han JM, Kim HL and Jung HJ: Ampelopsin inhibits cell proliferation and induces apoptosis in HL60 and K562 leukemia cells by downregulating AKT and NF-κB signaling pathways. Int J Mol Sci. 22(4265)2021.PubMed/NCBI View Article : Google Scholar | |
|
Chang H, Peng X, Bai Q, Zhou Y, Yu X, Zhang Q, Zhu J and Mi M: Ampelopsin suppresses breast carcinogenesis by inhibiting the mTOR signalling pathway. Carcinogenesis. 35:1847–1854. 2014.PubMed/NCBI View Article : Google Scholar | |
|
Lu CJ, He YF, Yuan WZ, Xiang LJ, Zhang J, Liang YR, Duan J, He YH and Li MY: Dihydromyricetin-mediated inhibition of the Notch1 pathway induces apoptosis in QGY7701 and HepG2 hepatoma cells. World J Gastroenterol. 23:6242–6251. 2017.PubMed/NCBI View Article : Google Scholar | |
|
Ye L, Yin G, Jiang M, Tu B, Li Z and Wang Y: Dihydromyricetin exhibits antitumor activity in nasopharyngeal cancer cell through antagonizing Wnt/β-catenin signaling. Integr Cancer Ther. 20(1534735421991217)2021.PubMed/NCBI View Article : Google Scholar | |
|
Zhang Q, Wang J, Zhang H and Zeng T: Dihydromyricetin inhibits oxidative stress and apoptosis in oxygen and glucose deprivation/reoxygenation-induced HT22 cells by activating the Nrf2/HO-1 pathway. Mol Med Rep. 23(397)2021.PubMed/NCBI View Article : Google Scholar | |
|
Park GB, Jeong JY and Kim D: Ampelopsin-induced reactive oxygen species enhance the apoptosis of colon cancer cells by activating endoplasmic reticulum stress-mediated AMPK/MAPK/XAF1 signaling. Oncol Lett. 14:7947–7956. 2017.PubMed/NCBI View Article : Google Scholar | |
|
Trepat X, Chen Z and Jacobson K: Cell migration. Compr Physiol. 2:2369–2392. 2012.PubMed/NCBI View Article : Google Scholar | |
|
Zanotelli MR, Zhang J and Reinhart-King CA: Mechanoresponsive metabolism in cancer cell migration and metastasis. Cell Metab. 33:1307–1321. 2021.PubMed/NCBI View Article : Google Scholar | |
|
Chen C, Xie L, Ren T, Huang Y, Xu J and Guo W: Immunotherapy for osteosarcoma: Fundamental mechanism, rationale, and recent breakthroughs. Cancer Lett. 500:1–10. 2021.PubMed/NCBI View Article : Google Scholar | |
|
Chou CH, Lu KH, Yang JS, Hsieh YH, Lin CW and Yang SF: Dihydromyricetin suppresses cell metastasis in human osteosarcoma through SP-1- and NF-κB-modulated urokinase plasminogen activator inhibition. Phytomedicine. 90(153642)2021.PubMed/NCBI View Article : Google Scholar | |
|
Wang K, Yang SF, Hsieh YH, Chang YY, Yu NY, Lin HW and Lin HY: Effects of dihydromyricetin on ARPE-19 cell migration through regulating matrix metalloproteinase-2 expression. Environ Toxicol. 33:1298–1303. 2018.PubMed/NCBI View Article : Google Scholar | |
|
Zhang QY, Li R, Zeng GF, Liu B, Liu J, Shu Y, Liu ZK, Qiu ZD, Wang DJ, Miao HL, et al: Dihydromyricetin inhibits migration and invasion of hepatoma cells through regulation of MMP-9 expression. World J Gastroenterol. 20:10082–10093. 2014.PubMed/NCBI View Article : Google Scholar | |
|
Liu T, Liu P, Ding F, Yu N, Li S, Wang S, Zhang X, Sun X, Chen Y, Wang F, et al: Ampelopsin reduces the migration and invasion of ovarian cancer cells via inhibition of epithelial-to-mesenchymal transition. Oncol Rep. 33:861–867. 2015.PubMed/NCBI View Article : Google Scholar | |
|
Wang FJ, Zong XY, DU JL, Wang WS, Yuan DP and Chen XB: [Effects of dihydromyricetin on the migration and invasion of human gastric cancer MKN45 cells and its mechanism]. Zhongguo Ying Yong Sheng Li Xue Za Zhi. 35:428–432. 2019.PubMed/NCBI View Article : Google Scholar : (In Chinese). | |
|
Zheng HQ and Liu DY: Anti-invasive and anti-metastatic effect of ampelopsin on melanoma. Ai Zheng. 22:363–367. 2003.PubMed/NCBIIn Chinese. | |
|
Huang CC, Su CW, Wang PH, Lu YT, Ho YT, Yang SF, Hsin CH and Lin CW: Dihydromyricetin inhibits cancer cell migration and matrix metalloproteinases-2 expression in human nasopharyngeal carcinoma through extracellular signal-regulated kinase signaling pathway. Environ Toxicol. 37:1244–1253. 2022.PubMed/NCBI View Article : Google Scholar | |
|
Chen L, Yang ZS, Zhou YZ, Deng Y, Jiang P and Tan SL: Dihydromyricetin inhibits cell proliferation, migration, invasion and promotes apoptosis via regulating miR-21 in human cholangiocarcinoma cells. J Cancer. 11:5689–5699. 2020.PubMed/NCBI View Article : Google Scholar | |
|
Sahoo BM, Banik BK, Borah P and Jain A: Reactive oxygen species (ROS): Key components in cancer therapies. Anticancer Agents Med Chem. 22:215–222. 2022.PubMed/NCBI View Article : Google Scholar | |
|
Villalpando-Rodriguez GE and Gibson SB: Reactive oxygen species (ROS) Regulates different types of cell death by acting as a rheostat. Oxid Med Cell Longev. 2021(9912436)2021.PubMed/NCBI View Article : Google Scholar | |
|
Kocaturk NM, Akkoc Y, Kig C, Bayraktar O, Gozuacik D and Kutlu O: Autophagy as a molecular target for cancer treatment. Eur J Pharm Sci. 134:116–137. 2019.PubMed/NCBI View Article : Google Scholar | |
|
Feng Y, He D, Yao Z and Klionsky DJ: The machinery of macroautophagy. Cell Res. 24:24–41. 2014.PubMed/NCBI View Article : Google Scholar | |
|
Onorati AV, Dyczynski M, Ojha R and Amaravadi RK: Targeting autophagy in cancer. Cancer. 124:3307–3318. 2018.PubMed/NCBI View Article : Google Scholar | |
|
Zhou Y, Shu F, Liang X, Chang H, Shi L, Peng X, Zhu J and Mi M: Ampelopsin induces cell growth inhibition and apoptosis in breast cancer cells through ROS generation and endoplasmic reticulum stress pathway. PLoS One. 9(e89021)2014.PubMed/NCBI View Article : Google Scholar | |
|
Zhou DZ, Sun HY, Yue JQ, Peng Y, Chen YM and Zhong ZJ: Dihydromyricetin induces apoptosis and cytoprotective autophagy through ROS-NF-κB signalling in human melanoma cells. Free Radic Res. 51:517–528. 2017.PubMed/NCBI View Article : Google Scholar | |
|
Liu B, Tan X, Liang J, Wu S, Liu J, Zhang Q and Zhu R: A reduction in reactive oxygen species contributes to dihydromyricetin-induced apoptosis in human hepatocellular carcinoma cells. Sci Rep. 4(7041)2014.PubMed/NCBI View Article : Google Scholar | |
|
Qi S, Xin Y, Guo Y, Diao Y, Kou X, Luo L and Yin Z: Ampelopsin reduces endotoxic inflammation via repressing ROS-mediated activation of PI3K/Akt/NF-κB signaling pathways. Int Immunopharmacol. 12:278–287. 2012.PubMed/NCBI View Article : Google Scholar | |
|
Tan M, Jiang B, Wang H, Ouyang W, Chen X, Wang T, Dong D, Yi S, Yi J, Huang Y, et al: Dihydromyricetin induced lncRNA MALAT1-TFEB-dependent autophagic cell death in cutaneous squamous cell carcinoma. J Cancer. 10:4245–4255. 2019.PubMed/NCBI View Article : Google Scholar | |
|
Tuli HS, Mistry H, Kaur G, Aggarwal D, Garg VK, Mittal S, Yerer MB, Sak K and Khan MA: Gallic Acid: A dietary polyphenol that exhibits anti-neoplastic activities by modulating multiple oncogenic targets. Anticancer Agents Med Chem. 22:499–514. 2022.PubMed/NCBI View Article : Google Scholar | |
|
Tuli HS, Kashyap D, Sharma AK and Sandhu SS: Molecular aspects of melatonin (MLT)-mediated therapeutic effects. Life Sci. 135:147–157. 2015.PubMed/NCBI View Article : Google Scholar | |
|
Manu KA, Shanmugam MK, Ramachandran L, Li F, Siveen KS, Chinnathambi A, Zayed ME, Alharbi SA, Arfuso F, Kumar AP, et al: Isorhamnetin augments the anti-tumor effect of capecitabine through the negative regulation of NF-κB signaling cascade in gastric cancer. Cancer Lett. 363:28–36. 2015.PubMed/NCBI View Article : Google Scholar | |
|
Manu KA, Shanmugam MK, Li F, Chen L, Siveen KS, Ahn KS, Kumar AP and Sethi G: Simvastatin sensitizes human gastric cancer xenograft in nude mice to capecitabine by suppressing nuclear factor-kappa B-regulated gene products. J Mol Med (Berl). 92:267–276. 2014.PubMed/NCBI View Article : Google Scholar | |
|
Jiang L, Zhang Q, Ren H, Ma S, Lu C, Liu B, Liu J, Liang J, Li M and Zhu R: Dihydromyricetin enhances the chemo-sensitivity of nedaplatin via regulation of the p53/Bcl-2 pathway in hepatocellular carcinoma cells. PLoS One. 10(e0124994)2015.PubMed/NCBI View Article : Google Scholar | |
|
He MH, Zhang Q, Shu G, Lin JC, Zhao L, Liang XX, Yin L, Shi F, Fu HL and Yuan ZX: Dihydromyricetin sensitizes human acute myeloid leukemia cells to retinoic acid-induced myeloid differentiation by activating STAT1. Biochem Biophys Res Commun. 495:1702–1707. 2018.PubMed/NCBI View Article : Google Scholar | |
|
Wang Z, Sun X, Feng Y, Liu X, Zhou L, Sui H, Ji Q, E Q, Chen J, Wu L and Li Q: Dihydromyricetin reverses MRP2-mediated MDR and enhances anticancer activity induced by oxaliplatin in colorectal cancer cells. Anticancer Drugs. 28:281–288. 2017.PubMed/NCBI View Article : Google Scholar | |
|
Wang Z, Sun X, Feng Y, Wang Y, Zhang L, Wang Y, Fang Z, Azami NLB, Sun M and Li Q: Dihydromyricetin reverses MRP2-induced multidrug resistance by preventing NF-κB-Nrf2 signaling in colorectal cancer cell. Phytomedicine. 82(153414)2021.PubMed/NCBI View Article : Google Scholar | |
|
Zhu XH, Lang HD, Wang XL, Hui SC, Zhou M, Kang C, Yi L, Mi MT and Zhang Y: Synergy between dihydromyricetin intervention and irinotecan chemotherapy delays the progression of colon cancer in mouse models. Food Funct. 10:2040–2049. 2019.PubMed/NCBI View Article : Google Scholar | |
|
Xu Y, Wang S, Chan HF, Lu H, Lin Z, He C and Chen M: Dihydromyricetin induces apoptosis and reverses drug resistance in ovarian cancer cells by p53-mediated Downregulation of Survivin. Sci Rep. 7(46060)2017.PubMed/NCBI View Article : Google Scholar | |
|
Wu M, Jiang M, Dong T, Xu L, Lv J, Xue M and Huang M: Reversal effect of dihydromyricetin on multiple drug resistance in SGC7901/5-FU cells. Asian Pac J Cancer Prev. 21:1269–1274. 2020.PubMed/NCBI View Article : Google Scholar | |
|
Dong S, Ji J, Hu L and Wang H: Dihydromyricetin alleviates acetaminophen-induced liver injury via the regulation of transformation, lipid homeostasis, cell death and regeneration. Life Sci. 227:20–29. 2019.PubMed/NCBI View Article : Google Scholar | |
|
Xiang D, Wang CG, Wang WQ, Shi CY, Xiong W, Wang MD and Fang JG: Gastrointestinal stability of dihydromyricetin, myricetin, and myricitrin: An in vitro investigation. Int J Food Sci Nutr. 68:704–711. 2017.PubMed/NCBI View Article : Google Scholar | |
|
Tong Q, Hou X, Fang J, Wang W, Xiong W, Liu X, Xie X and Shi C: Determination of dihydromyricetin in rat plasma by LC-MS/MS and its application to a pharmacokinetic study. J Pharm Biomed Anal. 114:455–461. 2015.PubMed/NCBI View Article : Google Scholar | |
|
Zhang R, Zhang H, Shi H, Zhang D, Zhang Z and Liu H: Strategic developments in the drug delivery of natural product dihydromyricetin: Applications, prospects, and challenges. Drug Deliv. 29:3052–3070. 2022.PubMed/NCBI View Article : Google Scholar | |
|
Chen S, Zhao X, Wan J, Ran L, Qin Y, Wang X, Gao Y, Shu F, Zhang Y, Liu P, et al: Dihydromyricetin improves glucose and lipid metabolism and exerts anti-inflammatory effects in nonalcoholic fatty liver disease: A randomized controlled trial. Pharmacol Res. 99:74–81. 2015.PubMed/NCBI View Article : Google Scholar | |
|
Ran L, Wang X, Lang H, Xu J, Wang J, Liu H, Mi M and Qin Y: Ampelopsis grossedentata supplementation effectively ameliorates the glycemic control in patients with type 2 diabetes mellitus. Eur J Clin Nutr. 73:776–782. 2019.PubMed/NCBI View Article : Google Scholar |









