



FOXP2 inhibits the aggressiveness of lung cancer cells by blocking TGFβ signaling
- Authors:
- Published online on: March 26, 2024 https://doi.org/10.3892/ol.2024.14361
- Article Number: 227
-
Copyright: © Su et al. This is an open access article distributed under the terms of Creative Commons Attribution License.
Abstract
Introduction
Lung cancer is one of the most common malignant tumors worldwide (1). The annual global incidence of lung cancer is over 1,800,000 (2). Various factors, such as smoking, habits and environment, contribute to the morbidity and mortality of lung cancer (3). In developed countries, lung cancer mortality has decreased since the 1990s (4). However, in China, patients with lung cancer have a higher mortality burden (5). Although great advances have been made in lung cancer treatment, such as chemotherapy, radiotherapy and surgery, the long-term overall survival rates remain unsatisfactory (6) due to limited access to diagnosis and treatment (7). Therefore, the identification of novel diagnostic markers for lung cancer is vital.
Forkhead box P2 (FOXP2), a transcription factor (8), is located on chromosome 7q31 and is a key regulator of metabolism, development and differentiation (9). FOXP2 is involved in embryonic and organ development, including that of the heart, lungs and central nervous system (10). Abnormal FOXP2 expression contributes to the pathogenesis of lung disorders, including lung cancer. For instance, FOXP2 alleviates LPS-induced apoptosis in human pulmonary alveolar epithelial cells and protects against acute lung injury (11). FOXP2 expression is decreased in patients with lung cancer (12). Notably, FOXP2-mediated upregulation of DNASE1L3 suppresses tumor cell proliferation and angiogenesis in lung adenocarcinoma (13). However, its role in lung cancer remains to be elucidated.
TGFβ regulates cell proliferation, migration and differentiation (14). However, TGFβ is frequently deregulated in carcinogenesis (15). TGFβ activation induced by tumor and stromal cells promotes tumor growth and metastasis (16). In addition, TGFβ signaling drives epithelial-to-mesenchymal transition processes (17), which may contribute to the chemoresistance and immune surveillance of tumor cells (18). Activated TGFβ ligand coordinates with its receptors to phosphorylate SMADs, which promotes nuclear translocation of SMADS to regulate the expression of the TGFβ target (19). In lung cancer, hypoxia-inducible factor 1-α-mediated activation of TGFβ/SMAD signaling accelerates tumor cell glycolysis and growth (20). Epigenetically stimulated TGFβ2 transcription enhances the radioresistance of lung cancer (21). However, the roles of TGFβ/SMAD signaling in lung cancer are still not fully understood.
The present study investigated the potential role of FOXP2 in lung cancer. Gene and protein expression were determined using RT-qPCR and western blotting, respectively. Functional analysis was performed using the CCK-8, colony formation, Transwell and TUNEL assays.
Materials and methods
Sampling
A total of 20 lung cancer tissues and adjacent tissues (>5 cm away from the tumor) [10 males and 10 females; 6 patients aged <60 years old (45–59 years old) and 14 patients aged ≥60 years old] were collected from patients hospitalized at Changzhou First People's Hospital (Jiangsu, China). The tissues were immediately frozen in liquid nitrogen and stored at −80°C for further processing. All diagnoses of non-small cell lung cancer were confirmed using pathological assays, including computed tomography, nuclear magnetic resonance imaging and immunohistochemistry. Patients who had previously received chemotherapy or radiotherapy were excluded from the present study. The present study was approved by the Ethics Committee of Changzhou First People's Hospital [Jiangsu, China; approval no. (2019)003].
Cell culture
Human lung cancer (A549, H1975 and H596) and human bronchial epithelial (16HBE) cell lines were provided by ATCC. Cells were cultured in RPMI-1640 medium (HyClone; Cytiva) containing 10% FBS at 37°C in an incubator (Thermo Fisher Scientific, Inc.) with 5% CO2. Cells were treated with 10 µM of SRI-011381 (MedChemExpress), an agonist of TGFβ signaling.
Cell transfection
pcDNA3.1 and pcDNA3.1-FOXP2 were obtained from Shanghai GenePharma Co., Ltd. A549 Cells with good growth state were taken for seed plate and transfected when the cell density reached 60%. The culture medium was replaced with a non-antibiotic medium 12 h before transfection. Cells were divided into the following groups: Control, untreated; pc-negative control (NC), transfected with 5 µl of Lipofectamine® 2000 (Invitrogen; Thermo Fisher Scientific, Inc.) and 20 µM pcDNA3.1 for 6 h at 37°C; pc-FOXP2, transfected with 5 µl of Lipofectamine® 2000 (Invitrogen; Thermo Fisher Scientific, Inc.) and 20 µM pcDNA3.1-FOXP2 for 6 h at 37°C and then replaced with complete medium and culture continued for 24–48 h.
Reverse transcription-quantitative (RT-q) PCR
All experimental operations were performed according to the manufacturer's protocols. When the cell density reached 1×106, total RNA was extracted from the cells. cDNA was synthesized using the RevertAid First Strand cDNA Synthesis Kit (cat. no. K1622; Thermo Fisher Scientific, Inc.). The mRNA expression was determined using PCR with the FastStart Universal SYBR Green Master (Rox) Kit (cat. no. 04913914001; Roche Diagnostics). mRNA expression was normalized to that of GAPDH. Results were measured using the 2−ΔΔCq method (22). The PCR conditions were as follows: Pre-denaturation at 95°C for 1 min, followed by denaturation at 95°C for 15 sec, and annealing and extension at 60°C for 30 sec, for 40 cycles. This was repeated three times for each set. The primer sequences were: GAPDH, F: 5′ AGAAGGTGGTGAAGCAGGCGTC 3′ and R: 5′ AAAGGTGGAGGAGTGGGTGTCG 3′; and FOXP2, F: 5′-GATGCAACAACTCCAGCAG-3′ and R: 5′-AGGACTTAAGCCAGCTTGAG-3′.
Western blotting
Cells in good condition were collected and the cell culture medium was discarded. Thereafter, cells were washed twice with PBS, RIPA lysis buffer (cat. no. BL504A; Biosharp Life Sciences) was added and the cells were shaken on ice for 15 min. The supernatant was centrifuged at 4°C, at 10,000 × g, for 5 min, and then the sample was heated at 100°C for 10 min. The protein concentration was determined using the BCA method. The protein (20 µg/lane) was isolated using SDS-PAGE (10%; 120 V) and transferred onto PVDF membranes (MilliporeSigma). Thereafter, the PVDF membranes were treated with 5% non-fat milk at room temperature to block them for 30 min, and incubated with primary antibodies at 4°C overnight: FOXP2 (cat. no. ab16046; 1:1,000), BAX (cat. no. ab32503; 1:2,500), BCL-2 (cat. no. ab182858; 1:2,000), p-SMAD3 (cat. no. ab52903; 1:2,000), SMAD3 (cat. no. ab208182; 1:1,000), SMAD4 (cat. no. ab40759; 1:5,000), TGFβR1 (cat. no. ab235578; 1:1,000), zinc finger E-box binding homeobox 1 (ZEB1; cat. no. ab203829; 1:500), zinc finger protein SNAI1 (SNAIL; cat. no. ab216347; 1:1,000) and GAPDH (cat. no. ab181602; 1:5,000) and then with HRP-labeled secondary antibody incubated at room temperature for 1 h. (cat. no. ab205718; 1:10,000). All antibodies were provided by Abcam. Proteins were visualized using an enhanced chemiluminescence kit (Tanon Science & Technology Co., Ltd.). Finally, ImageJ (National Institutes of Health) was used to analyze the gray value of the images.
CCK-8 assay
The cells were plated in 24-well plates at a density of 1×105 cells per well and incubated for 0, 24, 48 and 72 h after transfection at 37°C. The cells were then cultured with CCK-8 (10 µl; Beijing Solarbio Science & Technology Co., Ltd.) and cultured for another 4 h at 37°C. Subsequently, absorbance values were determined using a microplate reader at a wavelength of 450 nm.
Colony formation assay
Cells were plated in a 96-well plate precoated with soft agar and cultured at 37°C in a 5% CO2 incubator for 14 days. After fixing with 100% methanol at room temperature for 30 min, cells were stained with 1% crystal violet at room temperature for 15 min. Images were captured under a microscope (Leica Microsystems GmbH). Colonies with diameters >2 mm in the predetermined fields of interest were counted (magnification, ×200).
Transwell assay
Transwell chambers were pre-coated with Matrigel (BD Biosciences) and placed on ice for 30 min to form an even coating. Homogeneous serum-free cell suspensions (5×105 cells/well) were added to the upper chambers and the lower chambers were supplemented with 10% FBS. Transwell culture dishes were placed in a 5% CO2 cell incubator at 37°C for 24 h. Cells in the lower chamber were fixed and stained with 1% crystal violet (Beyotime Institute of Biotechnology). Finally, the number of migrated or invaded cells in predetermined fields of interest was calculated based on images captured using a microscope (CKX53; Olympus Corporation; magnification, ×400).
TUNEL assay
Cells were harvested, fixed in 4% paraformaldehyde (MilliporeSigma) at room temperature for 15 min and permeabilized with 0.25% Triton-X100 (Dalian Meilun Biotechnology Co., Ltd.). Thereafter, cells were stained using an in situ cell death detection kit (MilliporeSigma). Images were visualized using a fluorescence microscope (Nikon Corporation). The cell death rate was calculated as TUNEL-positive cells/total cells ×100.
Statistical analysis
Statistical analyses were performed using GraphPad software, version 9.5.1 (GraphPad; Dotmatics). Data were presented as mean ± standard deviation. Student's t-test was performed to analyze the differences between two groups, whereas one-way analysis of variance and Tukey's post hoc test were applied for multigroup analysis. P<0.05 was considered to indicate a statistically significant difference.
Results
FOXP2 is downregulated in lung cancer
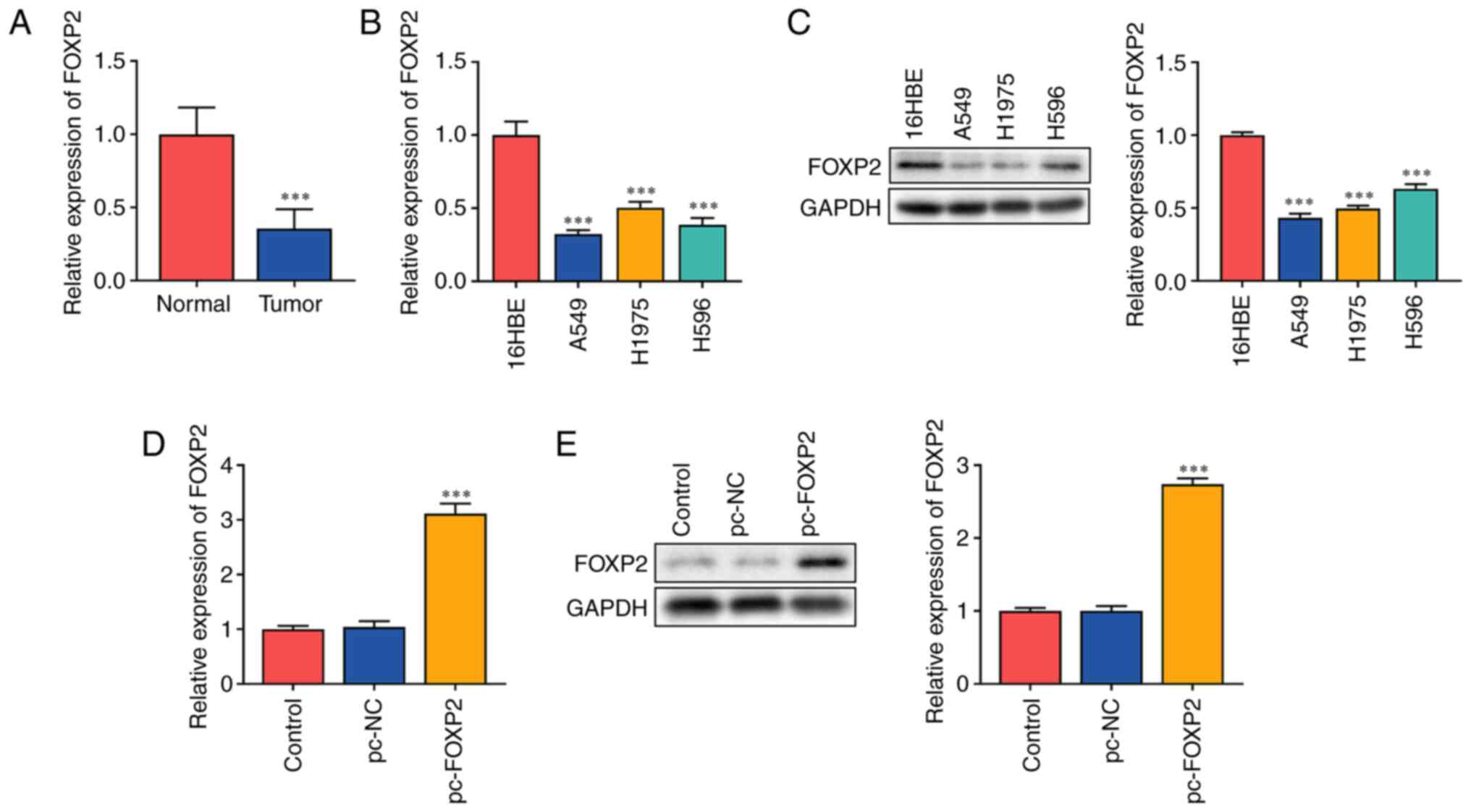
The present study hypothesized that FOXP2 functions as an antitumor gene in lung cancer. It was found that FOXP2 mRNA expression significantly decreased in patients with lung cancer (Fig. 1A). In addition, mRNA (Fig. 1B) and protein (Fig. 1C) expressions of FOXP2 were markedly decreased in lung cancer cells. A549 cells with a significant difference in FOXP2 expression were used for subsequent experiments. The potential role of FOXP2 in lung cancer was further investigated. A549 cells were transfected with FOXP2 overexpression plasmids. FOXP2 expression in the pc-FOXP2 group was significantly increased at both the mRNA and protein levels (Fig. 1D and E), suggesting that the cells were successfully transfected.
FOXP2 overexpression suppresses the aggressiveness of A549 cells
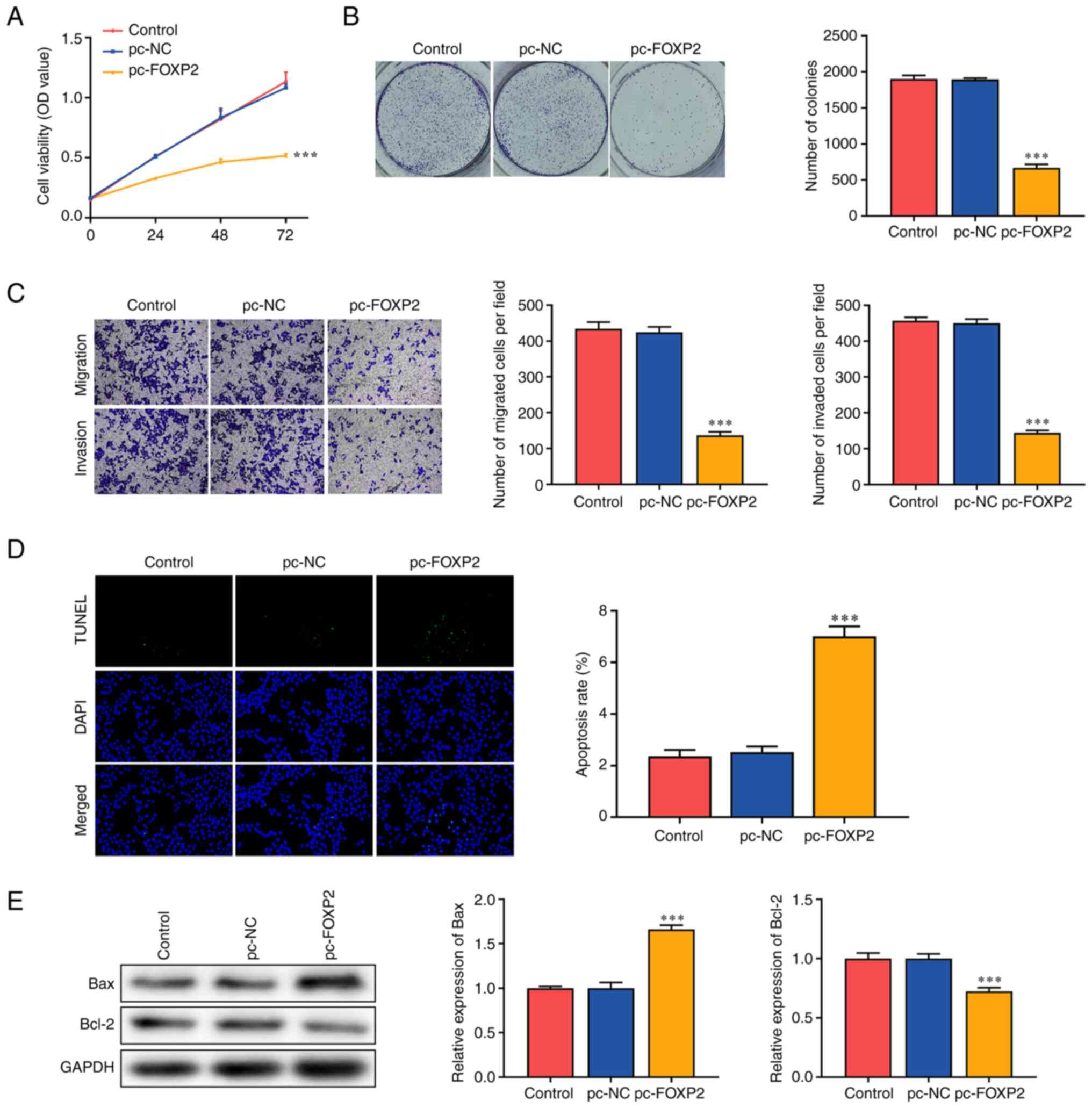
Functional assays were performed to determine the effects of FOXP2 on lung cancer cell function. The overexpression of FOXP2 significantly suppressed the viability of A549 cells compared with that in the pc-NC group (Fig. 2A). In addition, FOXP2 overexpression markedly inhibited colony formation in A549 cells (Fig. 2B). Migratory and invasive abilities were significantly suppressed in the pc-FOXP2 group (Fig. 2C). FOXP2 overexpression significantly increased the TUNEL-positive cells (Fig. 2D). In addition, FOXP2 overexpression increased BAX expression and suppressed BCL-2 expression (Fig. 2E). These findings suggested that FOXP2 overexpression suppressed the malignant behavior of lung cancer cells.
FOXP2 blocks TGFβ/SMAD signaling in lung cancer
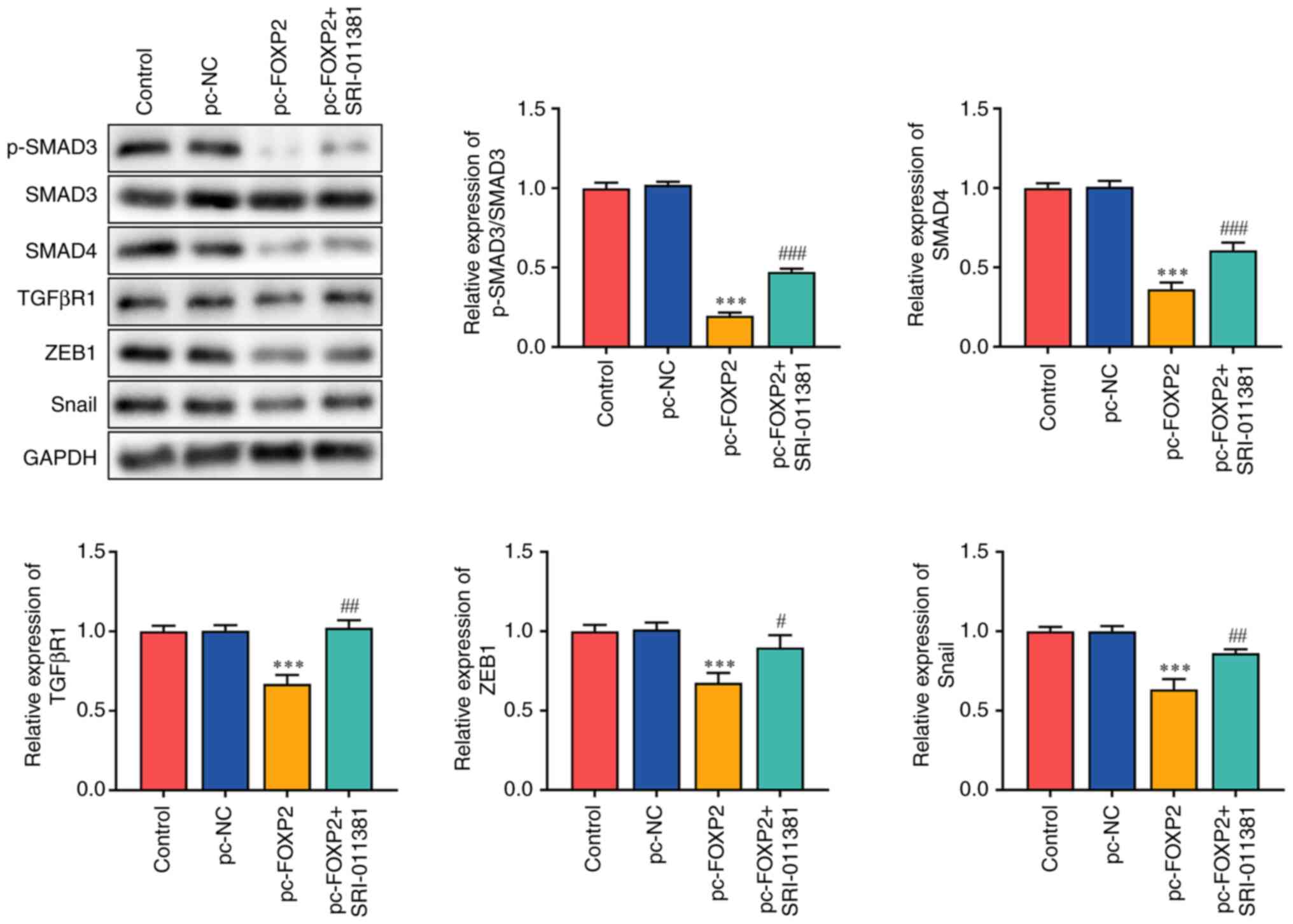
TGFβ/SMAD is involved in the carcinogenesis of lung cancer (23–25). FOXP2 overexpression suppresses tumor aggressiveness. Therefore, it was hypothesized that FOXP2 may inhibit the development of lung cancer by targeting TGFβ signaling. It was found that FOXP2 overexpression significantly inhibited SMAD3 phosphorylation and suppressed the protein expression of SMAD4, TGFβR1, ZEB1 and SNAIL. However, the TGFβ/SMAD signaling agonist SRI-011381 reversed the effects of pc-FOXP2 (Fig. 3).
Activation of TGFβ/SMAD signaling promotes the malignant behaviors of lung cancer
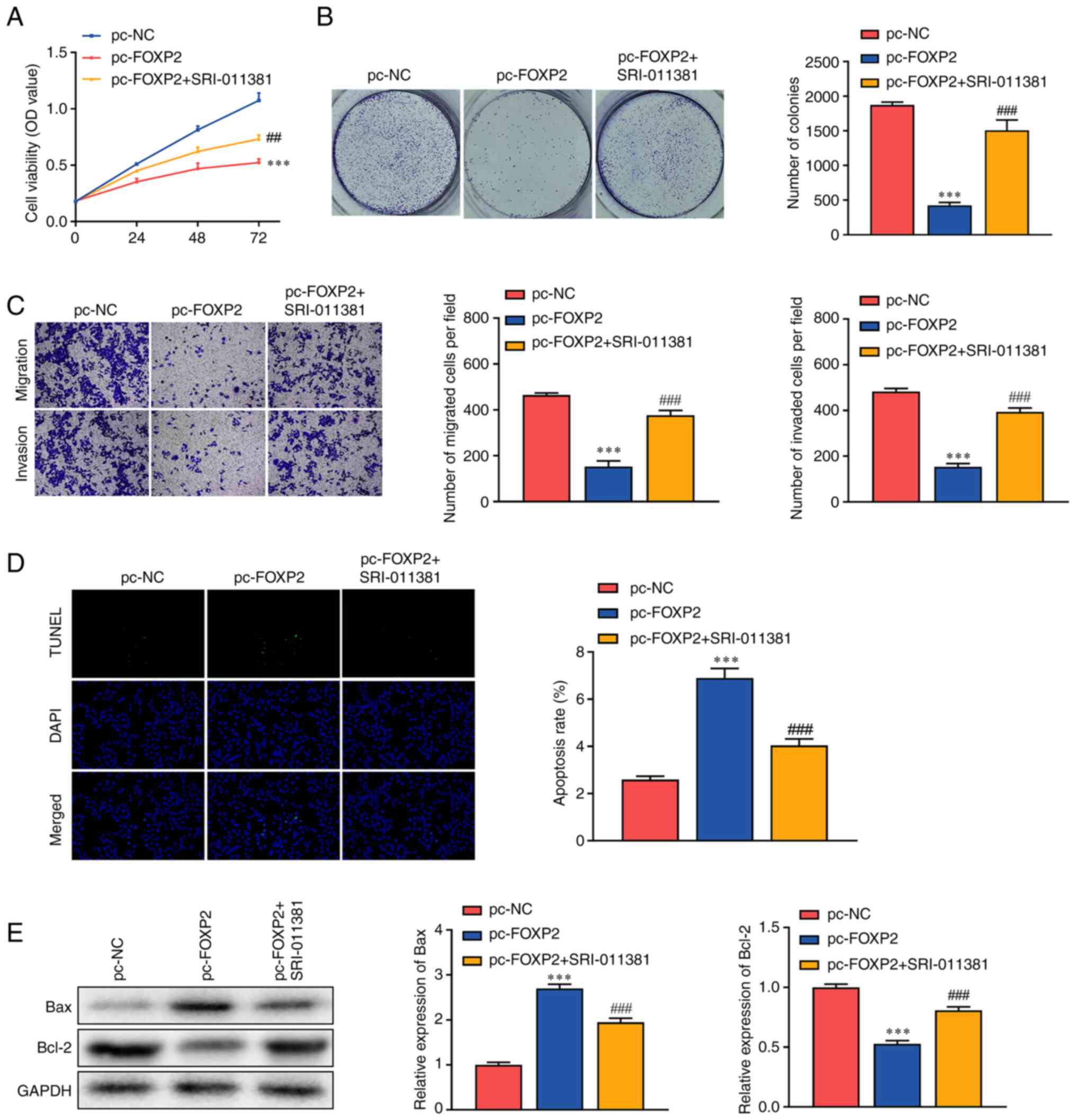
Rescue assays were conducted to verify the role of FOXP2 and TGFβ/SMAD in lung cancer. Following exposure to SRI-011381, a TGFβ/SMAD signaling agonist, the inhibition of cell viability induced by FOXP2 overexpression was alleviated (Fig. 4A). In addition, SRI-011381 treatment significantly abrogated the effects of FOXP2 overexpression and increased the number of tumor cell clones (Fig. 4B). This was consistent with the results of the Transwell assay. In addition, the inhibition of tumor cell migration and invasion induced by FOXP2 overexpression was markedly abrogated by SRI-011381 (Fig. 4C). In addition, SRI-011381 treatment significantly alleviated the effects of FOXP2 overexpression and suppressed apoptosis in A549 cells (Fig. 4D). SRI-011381 treatment significantly dampened the effects of FOXP2 overexpression, increased BCL-2 protein expression and downregulated BAX. These findings suggested that FOXP2 may suppress the aggressiveness of lung cancer by targeting TGFβ/SMAD signaling.
Discussion
In the present study, FOXP2 expression was downregulated in lung cancer. Notably, FOXP2 overexpression suppressed the proliferative, migratory and invasive abilities of lung cancer cells and promoted tumor cell apoptosis. In addition, FOXP2 blocked TGFβ signaling, the activation of which enhances malignant behaviors in tumor cells.
Increasing evidence indicates that FOXP2 functions as an oncogene in various types of cancer. For instance, circST3GAL6-mediates upregulation of FOXP2 promotes apoptosis and autophagy in gastric cancer (26). FOXP2 overexpression inhibits the migration of colon cancer (27). However, the role of FOXP2 in cancer remains unclear. Activation of HN1L/FOXP2 signaling-mediated stemness promotes tumor growth and migration in prostate cancer (28). In addition, FOXP2 overexpression promotes the migration and invasion of colorectal cancer cells (29), suggesting that it may also function as an oncogene. Therefore, identifying the exact role of FOXP2 in lung cancer is vital. In the present study, FOXP2 expression was decreased in lung cancer cells. In addition, FOXP2 overexpression suppressed the proliferative, migratory and invasive abilities of lung cancer cells, suggesting that FOXP2 may function as an antitumor gene in lung cancer. These findings were consistent with those of previous studies (12,13).
FOXP2 alters cellular functions by regulating the expression of its targets (30). For instance, FOXP2 epigenetically activates RPS6KA6 to enhance tumor cell apoptosis in thyroid cancer (31). FOXP2 interacts with caspase-1 to drive tumor cell pyroptosis in colorectal cancer (23). In the present study, FOXP2 blocked TGFβ signaling, which plays a key role in the pathogenesis of cancers. However, its role varies with the stages of tumors. At the early stages, TGFβ signaling functions as a tumor suppressor and promotes cell cycle arrest (24). However, the enrichment of proinflammatory TGFβ induces the degradation of epithelial functions and the acquisition of mesenchymal features (25), promoting tumor cell migration and invasion. In addition, the continuous release of TGFβ contributes to the immune evasion of tumor cells by recruiting macrophages, cancer-associated fibroblasts and neutrophils (32). In the present study, SRI-011381-mediated activation of TGFβ/SMAD signaling promoted the proliferation, migration and invasion of lung cancer cells and suppressed tumor cell apoptosis. These findings suggested that FOXP2 suppressed the aggressiveness of lung cancer cells by targeting TGFβ/SMAD signaling.
The present study had some limitations. First, it included only 20 participants and did not distinguish the malignant degree of lung cancer. Future studies with a larger sample size are needed to confirm the results and further studies are needed to investigate the correlation between FOXP2 expression and malignant degree of lung cancer. Second, avoiding recollection bias when obtaining past information was difficult. In addition, some of the clinical data were missing. Therefore, well-designed studies are warranted in the future.
In conclusion, FOXP2 functions as an antitumor gene in lung cancer. FOXP2 suppressed the proliferation, migration and invasion of lung cancer cells and promoted apoptosis in lung cancer cells by blocking TGFβ/SMAD signaling. Therefore, FOXP2/TGFβ/SMAD signaling may be a potential target for lung cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by Changzhou Key Medical Discipline (grant no. CZXK202205).
Availability of data and materials
The data generated in the present study may be requested from the corresponding author.
Authors' contributions
ZYL conceived and designed the study. WYS, SH and LZ performed the literature search, performed the experiments and data extraction. HB analyzed and interpreted the data. WYS and HB drafted the manuscript. ZYL, WYS and SH confirm the authenticity of all the raw data and revised the final version of the manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to participate
The present study was approved by the Ethics Committee of Changzhou First People's Hospital [Jiangsu, China; approval no. (2019)003]. All patients signed informed consent forms.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no conflicts of interest.
References
|
Hamann HA, Ver Hoeve ES, Carter-Harris L, Studts JL and Ostroff JS: Multilevel opportunities to address lung cancer stigma across the cancer control continuum. J Thorac Oncol. 13:1062–1075. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Zhuo C, Zhuang H, Gao X and Triplett PT: Lung cancer incidence in patients with schizophrenia: Meta-analysis. Br J Psychiatry. 215:704–711. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Nooreldeen R and Bach H: Current and Future development in lung cancer diagnosis. Int J Mol Sci. 22:86612021. View Article : Google Scholar : PubMed/NCBI | |
|
Zhou M, Wang H, Zeng X, Yin P, Zhu J, Chen W, Li X, Wang L, Wang L, Liu Y, et al: Mortality, morbidity, and risk factors in China and its provinces, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet. 394:1145–1158. 2019. View Article : Google Scholar : PubMed/NCBI | |
|
Yang D, Liu Y, Bai C, Wang X and Powell CA: Epidemiology of lung cancer and lung cancer screening programs in China and the United States. Cancer Lett. 468:82–87. 2020. View Article : Google Scholar : PubMed/NCBI | |
|
Wang M, Herbst RS and Boshoff C: Toward personalized treatment approaches for non-small-cell lung cancer. Nat Med. 27:1345–1356. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Zugazagoitia J and Paz-Ares L: Extensive-Stage small-cell lung cancer: First-Line and second-line treatment options. J Clin Oncol. 40:671–680. 2022. View Article : Google Scholar : PubMed/NCBI | |
|
Nudel R and Newbury DF: Foxp2. Wiley Interdiscip Rev Cogn Sci. 4:547–560. 2013. View Article : Google Scholar : PubMed/NCBI | |
|
den Hoed J, Devaraju K and Fisher SE: Molecular networks of the FOXP2 transcription factor in the brain. EMBO Rep. 22:e528032021. View Article : Google Scholar : PubMed/NCBI | |
|
Furlong R: FOXP2 tells a cautionary tale. Nat Rev Genet. 19:592–593. 2018. View Article : Google Scholar : PubMed/NCBI | |
|
Nan CC, Zhang N, Cheung KCP, Zhang HD, Li W, Hong CY, Chen HS, Liu XY, Li N and Cheng L: Knockdown of lncRNA MALAT1 Alleviates LPS-Induced acute lung injury via inhibiting apoptosis through the miR-194-5p/FOXP2 Axis. Front Cell Dev Biol. 8:5868692020. View Article : Google Scholar : PubMed/NCBI | |
|
Ren T, Liu C, Hou J and Shan F: Hsa_circ_0043265 suppresses proliferation, metastasis, EMT and promotes apoptosis in non-small cell lung cancer through miR-25-3p/FOXP2 pathway. Onco Targets Ther. 13:3867–3880. 2020. View Article : Google Scholar : PubMed/NCBI | |
|
Meng F, Yang X and Xiao P: DNASE1L3 regulation by transcription factor FOXP2 affects the proliferation, migration, invasion and tube formation of lung adenocarcinoma. Exp Ther Med. 25:722022. View Article : Google Scholar : PubMed/NCBI | |
|
Peng D, Fu M, Wang M, Wei Y and Wei X: Targeting TGF-β signal transduction for fibrosis and cancer therapy. Mol Cancer. 21:1042022. View Article : Google Scholar : PubMed/NCBI | |
|
Hao Y, Baker D and Ten Dijke P: TGF-beta-mediated epithelial-mesenchymal transition and cancer metastasis. Int J Mol Sci. 20:27672019. View Article : Google Scholar : PubMed/NCBI | |
|
Syed V. TGF-beta signaling in cancer. J Cell Biochem. 117:1279–1287. 2016. View Article : Google Scholar : PubMed/NCBI | |
|
Mortezaee K, Majidpoor J and Kharazinejad E: Epithelial-mesenchymal transition in cancer stemness and heterogeneity: Updated. Med Oncol. 39:1932022. View Article : Google Scholar : PubMed/NCBI | |
|
Giannelli G, Koudelkova P, Dituri F and Mikulits W: Role of epithelial to mesenchymal transition in hepatocellular carcinoma. J Hepatol. 65:798–808. 2016. View Article : Google Scholar : PubMed/NCBI | |
|
Zhou JY, Zheng SR, Liu J, Shi R, Yu HL and Wei M: MiR-519d facilitates the progression and metastasis of cervical cancer through direct targeting Smad7. Cancer Cell Int. 16:212016. View Article : Google Scholar : PubMed/NCBI | |
|
Huang Y, Chen Z, Lu T, Bi G, Li M, Liang J, Hu Z, Zheng Y, Yin J, Xi J, et al: HIF-1alpha switches the functionality of TGF-β signaling via changing the partners of smads to drive glucose metabolic reprogramming in non-small cell lung cancer. J Exp Clin Cancer Res. 40:3982021. View Article : Google Scholar : PubMed/NCBI | |
|
Jie X, Fong WP, Zhou R, Zhao Y, Zhao Y, Meng R, Zhang S, Dong X, Zhang T, Yang K, et al: USP9X-mediated KDM4C deubiquitination promotes lung cancer radioresistance by epigenetically inducing TGF-β2 transcription. Cell Death Differ. 28:2095–2111. 2021. View Article : Google Scholar : PubMed/NCBI | |
|
Livak KJ and Schmittgen TD: Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI | |
|
Liao P, Huang WH, Cao L, Wang T and Chen LM: Low expression of FOXP2 predicts poor survival and targets caspase-1 to inhibit cell pyroptosis in colorectal cancer. J Cancer. 13:1181–1192. 2022. View Article : Google Scholar : PubMed/NCBI | |
|
Colak S and Ten Dijke P: Targeting TGF-β signaling in cancer. Trends Cancer. 3:56–71. 2017. View Article : Google Scholar : PubMed/NCBI | |
|
Lee JH and Massague J: TGF-beta in developmental and fibrogenic EMTs. Semin Cancer Biol. 86((Pt 2)): 136–145. 2022. View Article : Google Scholar : PubMed/NCBI | |
|
Xu P, Zhang X, Cao J, Yang J, Chen Z, Wang W, Wang S, Zhang L, Xie L, Fang L, et al: The novel role of circular RNA ST3GAL6 on blocking gastric cancer malignant behaviours through autophagy regulated by the FOXP2/MET/mTOR axis. Clin Transl Med. 12:e7072022. View Article : Google Scholar : PubMed/NCBI | |
|
Zhao Q, Liu C, Cui Q, Luan X, Wang Q and Zhou C: miR-190b promotes colorectal cancer progression through targeting forkhead box protein P2. Exp Ther Med. 19:79–84. 2020.PubMed/NCBI | |
|
Nong S, Wang Z, Wei Z, Ma L, Guan Y and Ni J: HN1L promotes stem cell-like properties by regulating TGF-β signaling pathway through targeting FOXP2 in prostate cancer. Cell Biol Int. 46:83–95. 2022. View Article : Google Scholar : PubMed/NCBI | |
|
Huang R, Xiang G, Duan X, Wang H, He K and Xiao J: MiR-132-3p inhibits proliferation, invasion and migration of colorectal cancer cells via down-regulating FOXP2 expression. Acta Biochim Pol. 69:371–377. 2022.PubMed/NCBI | |
|
Vernes SC, Newbury DF, Abrahams BS, Winchester L, Nicod J, Groszer M, Alarcón M, Oliver PL, Davies KE, Geschwind DH, et al: A functional genetic link between distinct developmental language disorders. N Engl J Med. 359:2337–2345. 2008. View Article : Google Scholar : PubMed/NCBI | |
|
Yang F, Xiao Z and Zhang S: FOXP2 regulates thyroid cancer cell proliferation and apoptosis via transcriptional activation of RPS6KA6. Exp Ther Med. 23:4342022. View Article : Google Scholar : PubMed/NCBI | |
|
Tauriello DVF, Sancho E and Batlle E: Overcoming TGFβ-mediated immune evasion in cancer. Nat Rev Cancer. 22:25–44. 2022. View Article : Google Scholar : PubMed/NCBI |









